Embed Size (px)
Citation preview

Oeeologia (Berl.) 9, 215--222 (1972) �9 by Springer-Verlag 1972
Sind einfache ZeitverzSgerungen die Ursachen fiir periodische Populationsschwankungen?
Vergle ich e x p e r i m e n t e l l e r U n t e r s u c h u n g e n an Brachionus calyciflorus Pa l l a s ( R o t a t o r i a ) m i t C o m p u t e r - S i m u l a t i o n e n
Udo I t a l b a e h und t te inz J i i rgen B u r k h a r d t *
Zoologisches Institut der Universit~t Miinchen
Eingegangen am29, Januar 1972
Are Simple T ime Lags Respons ib le for Cyclic Var ia t ion of Popu la t ion Dens i ty ?
A Comparison of Laboratory Population Dynamics of Brachionus calyci]lorus Pallas (Rotatoria) with Computer Simulations
Summary. Laboratory populations of the rotifer Brachionus calyciflorus were cultured at different temperatures (25, 20, 15~ but otherwise at constant condi- tions. The population densities showed relatively constant oscillations (Figs. 1 to 3A-C). Amplitudes and frequencies of the oscillations were positively correlated with temperature (Table 1). A test was made, whether the logistic growth function with simple time lag is able to describe the population curves. There arc strong similarities between the simulations (Figs. 1--3 E) and the real population dynamics if minor adjustments of the empirically determined parameters are made. There- fore it is suggested that time lags are responsible for the observed oscillations. However, the actual time lags probably do not act in the simple manner of the model, because birth and death rates react with different time lags, and both parameters are dependent on individual age and population density. A more com- plex mode], which incorporates these modifications, should lead to a more realistic description of the observed oscillations.
Einlei tung
Schwankungen der Ind iv iduend ich te t icr ischer Popu la t ionen shad vor a l lem auf Ver/~nderungen des Milieus (z. B. W i t t e r u n g oder Nahrungs- angebot ) zuriickzuffihren. Nich t sel ten beobach te t man aber auch un te r kons t an t en Umwel tbed ingungen eine Var ia t ion dcr Ind iv iducnd ich te . So f indet m a n zykl isehe F l u k t u a t i o n e n ha Laborpopu la t i onen des R/~der- t ieres Brachionus calyci/lorus, die un te r kons t a n t e n Bedingungcn gehal-
* W i r danken Herrn Prof. J. Jaeobs nnd Herin Dr. It. Kaiser fiir die kritische Durchsicht des Manuskripts.

216 U. Halbach und H. J. ]~urkhardt:
ten werden (Abb. 1--3, jeweils A--C). Es wurde postuliert (Halbach, 1969, 1970), dab ZeitverzSgerungen ffir diese Oszillationen der Tierdichte verantwortlieh sind: Mit zunehmender Tierdiehte ~ r d die Nahrungs- menge fiir jedes einzelne Tier geringer; Hunger ffihrt jedoch zu einer Herabsetzung der Lebensdauer und der Reproduktion (Halbaeh, 1971 a). Mit Hflfe dieses Meehanismus kann die Tierdichte der zur Verffigung stehenden Nahrung angeglichen werden. Die Anpassung an die momen- tane Nahrungssituation erfolgt jedoeh nicht unmittelbar, sondern mit einer zeitlichen Verz6gerung, da zwischen der Nahrungsaufnahme und dem Umsatz der Nahrung in Nachkommen notwendigerweise eine gewisse Zeitspanne liegt. Sie ist die Ursache daffir, dal] die waehsende Population fiber die Gleiehgewiehtsdichte hinaussehieBt. Die Folge sind Oszfllationen um die Gleiehgewiehtsdiehte oder Kapaziti~t.
Hutchinson (1948) hat ffir selbstregulierende Populationen die modi- fizierte logistisehe Gleichung mit ZeitverzSgerung vorgeschlagen:
dN(t) ~ ~,'f [K--N(t-T)] dt - . '~ , ( t )[ ~ ], (1)
wobei t =Zeit , N = Individuendiehte, K---- Kapazit~t, T---- Zeitverz5ge- rung. Diese Gleichung ist yon Cunningham (1954) und yon Wangersky und Cunningham (1956, 1957) genauer analysiert worden. Naeh dieser Analyse kann man bei bestindigen Oszillationen die ZeitverzSgerung T naeh folgender Beziehung aus der Frequenz ] berechnen:
1 T = ].~.V~. (2)
Es soll hier untersucht werden, wieweit dieses Modell in der Lage ist, die experimentell ermittelten Populationskurven yon Brachionus calyci- /lotus zu beschreiben.
Material und Methode
A. Tiermaterial und Versuchslculturen
Bei den verwendeten Labortieren yon Brachionus calyci/lorus han- delt es sich um einen Klon. Das amiktische (parthenogenetisehe) Weib- ehen, yon dem dieser Klon abstammt, war im Mai 1966 aus dem Wolfsee bei Seheinfeld (Steigerwald) isoliert worden.
Die Haltung erfolgte in synthetisehem Sfil~wasser in Petrisehalen yon 10 nil Inhalt. Die Tiere warden t~glieh mit Haarpipetten in frisches Medium umgesetzt, wobei ihre Dichte bestimmt wurde. Als Futter erhielten sie tiglieh 2 • l0 s Chlorella vulgaris/ml aus Flfissigkeitskulturen.

Simulation oszillierender Populationen 217
Die Versuchskulturen wurden bei 25, 20 und 15~ (jewefls ~: I~ im Dauerdunkel gehalten. Weitere Ehlzelheiten zur Methodik linden sich bei Halbach (1971b).
B. Computer-Simulationen
Zur zeitkontinuierlichen digitalen Simulation des logistischen Modells mit Zeitverz6gerung [G1. (1)] warde die Simulationssprache D SL/90 ver- wendet (Syn and Wyman, 1965; Burkhardt, 1970). Die Berechnungen wurden auf der IBM 7094 des Deutschen Rechenzentrums Darmstadt durchgeffihrt. Zur numerischen Integration wurden Runge-Kutta-Ver- fahren mit variabler Schrittwelte verwendet. Die Integration erfolgte jeweils fiber 60 Tage.
Ergebnisse
A. Experimental- Populationen
Es wurden jeweils 3 Versuchspopulationen bei 25, 20 und 15~ ge- halten. Der Verlauf ihrer Populationsdiehten ist in Abb. 1--3 (jewefls A--C) wiedergegeben. Mit abnehmender Temperatur verlieren die Oszfl- lationen an Heftigkeit, d.h. Amplitude und Frequenz nehmen ab. Die potentielle Wachstumsrate, d.i. die Waehstumsrate w~hrend des ex- ponentiellen Wachstums, f/~llt eberffalls mit der Temperatur ab, wi~hrend die Kapazitiit, d.i. die mittlere Tierdichte im Gleichgewieht, mit sinken- der Temperatur zunimmt (Tabelle 1).
Tabelle 1. Potentielle Wachstumsrate (r), Kapazit~t (K), Frequenz ([) und Ampli- tude (A) der Versuchspopulationen in Abh~ngigkeit yon der Temperatm" (Streuungs-
real3 = Standardabweichung)
15 ~ C 20 ~ C 25 ~ C
PotentieIIe Wachstumsr~te r (Nachkommen/~ • Std) 0,34 • 0,06 0,48 • 0,07 0,82 ~= 0,07
Kapazit~t K (Tiere/ml) 74,7 66,8 57,3
Frequenz ] (Oszillationen/Tag) 0,05 0,09 0,12
Amplitude A (Tiere/ml) 12,1 31,6 58,0
ZeitverzSgerung T (Tage) 4,5 2,5 1,9
218 U. Halbach und H. J. Burkhardt:
150
50
150 F
~176 A |
~ loo
._~
50
, i f i J i ,
150
50
. . . . . r . . i , , r , ~
0 10 20 30
Tage
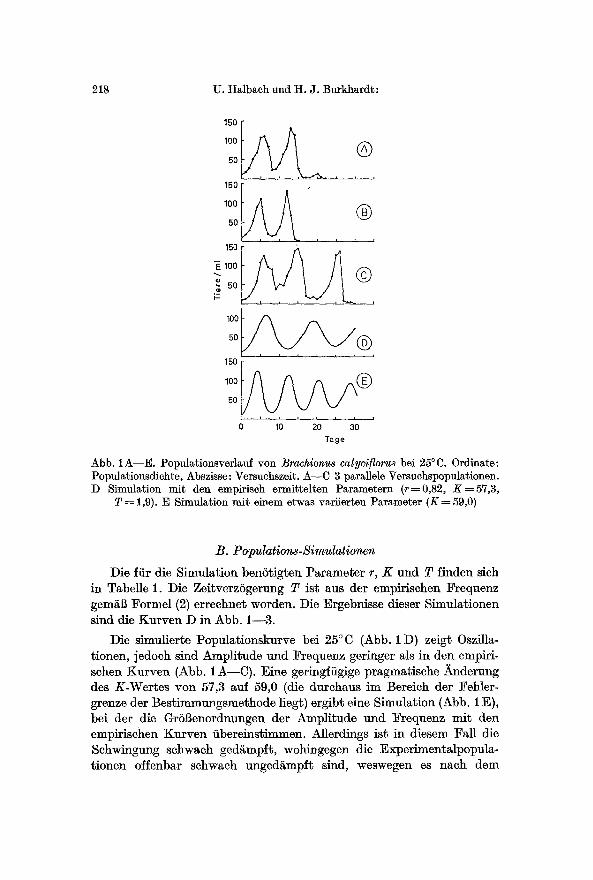
Abb. 1A--E. Populationsverlauf yon Brachionus calyci]lorus bei 25~ Ordinate: Populationsdichte, Abszisse: Versuchszeit. A--C 3 parallele Versuchspopulationen. D Simulation mit den empirisch ermittelten Parametern (r----0,82, K=57,3,
T ~ 1,9). E Simulation mit einem etwas variierten Parameter (K ~ 59,0)
B. Populations-Simulationen
Die fiir die Simulation benStigten Parameter r, K und T linden sich in Tabelle 1. Die ZeitverzSgerung T ist aus der empirischen Frequenz gem~B Formel (2) erreehnet worden. Die Ergebnisse dieser Simulationen sind die Kurven D in Abb. 1--3.
Die simulierte Populationskurve bei 25~ (Abb. 1 D) zeigt Oszilla- tionen, jedoch sind Amplitude uud Frequenz geringer als in den empiri- schen Kurven (Abb. 1 A---C). Eine geringfiigige pragmatische ~nderung des K-Wertes yon 57,3 auf 59,0 (die durchaus im Bereich der Fehler- grenze der Bestimmungsmethode liegt) ergibt eine Simulation (Abb. 1 E), bei der die GrS•enordnungen der Amplitude und Frequenz mit den empirischen Knrven fibereinstimmen. Allerdings ist in diesem Fall die Schwingung schwach ged~mpft, wohingegen die Experimentalpopula. tionen offenbar schwach unged~mpft sind, weswegen es nach dem
Simulation oszillbrender Populationen 219
100
50
100
50
150
100
~ 5o
* 100 1 -
50
150
100
50
0
, , , l l , , . l l r
. r . r l . r l r ~ f f ~
10 20 30 40 50 60 Tage
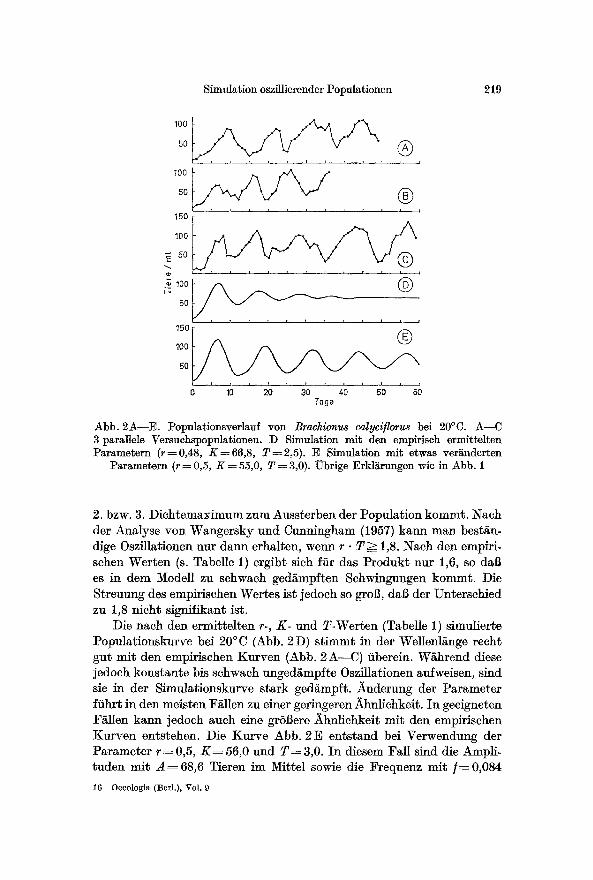
Abb. 2A--E. Populationsverlauf yon Brachionus calyviflorus bei 20~ A--C 3 parallele Versuchspopulationen. D Simulation mit den empLrisch ermittelten Parametern (r=0,48, K = 66,8, T=2,5). E Simulation mit etwas ver~nderten
Parametern (r = 0,5, K ~ 55,0, T = 3,0). Obrige Erklgrungen wie in Abb. 1
2. bzw. 3. Dichtemaximum zum Aussterben der Population komm~. Nach der Analyse yon Wangersky und Cunningham (1957) kann man best~n- dige Oszflla*ionen nur dann erhalten, wenn r �9 T ~ 1,8. Nach den empiri- schen Werten (s. Tabelle 1) ergib~ sich ffir das Produk* nut 1,6, so dab es in dem Modell zu schwach geds Schwingungen kommt. Die Streuung des empirischen Wertes ist jedoch so groB, dab der Unterschied zu 1,8 nicht signifik~nt ist.
Die nach den ermittelten r-, K- und T-Werten (Tabelle 1) simulierte Populationskurve bei 20~ (Abb. 2 D) s t immt in der Wellenl~nge recht gut mit den empirischen Kurven (Abb. 2A--C) fiberein. Wghrend diese jedoch konstante bis schwach ungedgmpfte Oszilla*ionen aufweisen, sind sie in der Simulationskurve stark ged~mpft. ~_uderung der Parameter ffihrt in den meisten Fgilen zu einer geringeren i~hnlichkei~. In geeigneten F~llen kann jedoch auch eine grSBere Ahnlichkeit mit den empirischen Kurven entstehen. Die Kurve Abb. 2E entstand bei Verwendung der Parameter r = 0,5, K = 56,0 und T : 3,0. In diesem Fail sind die Ampli- tuden mit A ~ 6 8 , 6 Tieren im Mittel sowie die Frequenz m i t / ~ 0 , 0 8 4
16 Oecologia (Berl.), Vol. 9
220 U. I-Ialbach und H. J. Burkhardt:
100[
100 [
50 ~ , 100 r
300 5O
T
1~176 I i , . i
10 20 30 40 Tclge
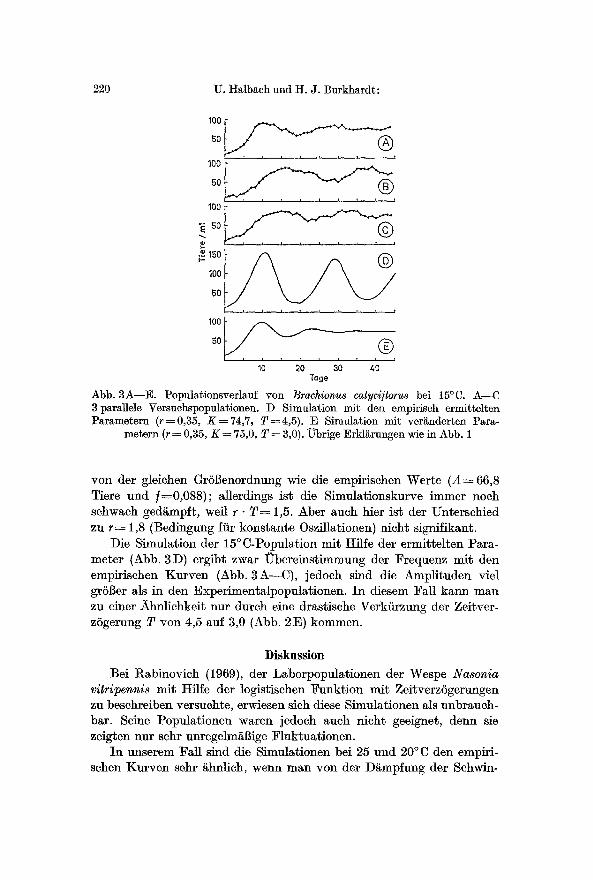
Abb. 3A--E. Populationsverlauf yon Brachionus calycl]loru8 bei 15~ A--C 3 parallele Versuchspopulationen. D Simulation mit den empiriseh ermitteltea Parametern (r=-0,35, K = 74,7, T=4,5). E Simulation mit ver~nderten Para-
metern (r = 0,35, K = 75,0, T = 3,0). ~brige Erkl~rungen wie in Abb. 1
yon der gleichen GrSl]enordnung wie die empirischen Werte ( A = 66,8 Tiere und f=0,088); allerdings ist die Simulationskurve immer noeh sehwaeh ged/~mpft, well r �9 T-= 1,5. Abet auch hier ist der Unterschied zu r = 1,8 (Bedingung fiir konstante 0szfllationen) nicht signifikant.
Die Simulation der 15~ mit Hflfe der ermittelten Para- meter (Abb. 3D) ergibt zwar ~bereinst immung der Frequenz mit den empirischen Kurven (Abb. 3A---C), jedoch sind die Amplituden vie1 grSl]er als in den Experimentalpopulationen. In diesem Fall kann man zu einer Xhnlichkeit nut dutch eine dras~ische Verkiirzung der Zeitver- zSgerung T yon 4,5 auf 3,0 (Abb. 2E) kommen.
Diskussion
Bei Rabinovich (1969), der Laborpopulationen der Wespe Nasonia vitripennis mit ttilfe der logistischen Funktion mit Zeitverz6gerungen zu beschreiben versuchte, erwiesen sich diese Simulationen als unbrauch- bar. Seine Populationen waren jedoch auch nicht geeignet, denn sie zeigten nur sehr unregelm/~Bige Fluktuationen.
In unserem Fall sind die Simulationen bei 25 und 20~ den empiri- schen Kurven sehr/ihnlich, wenn man yon der D~mpfung der Schwin-

Simulation oszillierender Populationen 221
gungen absieht. Die Unterschiede k6nnte man durch eine ungenaue Bestimmung der Zeitverz6gerung T [aus der Frequenz der empirischen Oszfllationen, vgl. G1. (2)] erkl~ren. Bei der Simulation der 15 ~ C-Popula- tion mug man - - um zu ~hnlichen Kurven zu kommen - - die Zeitver- z6gerung jedoch in einer Weise verkfirzen, die sich nicht ohne weiteres ans der Ungenauigkeit der Bestimmung erkls l~l~t.
Wenn auch die Obereinstimmung der Simula~ionen mit den empiri- schen Populationsdynamiken nur unvollkommen is~, bestgrken sie doch erheblich die Itypothese, dab Zeitverz6gerungen fiir die beobachteten Oszfllationen veranr sind, zumal es zur Zeit keine andere Erkl~- rung mit Khnlieher l~berzeugungskraft gibt. Es ist jedoch anzunehmen, dab die Zeitverz6gerungen nicht in so einfacher Weise wirken wie in dem vorliegenden Modell, sondern dal~ sie komplexer sind. Experimentelle Untersuchungen haben niimlich gezeigt, dal3 die Geburtsrate mit einer kfirzeren Zeitverz6gerung auf Anderungen der Dichte reagiert als die Sterberate (Seitz und Halbach, in Vorbereitung). Eine Untersuehung, wieweit diese Zeitverz6gerungen auch noch alters- und diehteabhi~ngig sind, steht noch aus. Bekannt ist jedoch, dab Geburts- und Sterberate eine Alters- und D ichteabh~ngigkeit besitzen. AuBerdem ist bekannt, dal~ der Dichteeinflu$ auf zwei verschiedene Weisen erfolgt: fiber die Abgabe sch~dlicher Metabolite in das Medium (Halbaeh, 1969) und fiber Nahrungskonkurrenz (ttalbach, 1971 a). Bei der Konstruktion eines rea- listischeren Modells sollten daher Alters- und Dichteabh~ngigkeiten berficksichtigt werden.
Literatur
Burkhardt, H. J. : Digitale Simulation zeitkontinuierlicher Systeme. Ver6ff. Dtsch. Rechenzentrum Darmstadt (1970).
Cunningbam, W. J. : A non-linear difference-differential equation of growth. Proc. nat. Acad. Sei. (Wash.) 40, 708--713 (1954).
Halbach, U. : Das Zusammenwirken yon Konkurrenz und R~iuber-Beute-Beziehun- fen bei R~dertieren. Zool. Anz., Suppl. 33, 72--79 (1969).
Halbaeh, U. : Einflul] der Temperatur auf die Populationsdynamik des planktischen R~dertieres Brachionus calyciflorus Pallas. Oecologia (Berl.) 4, 176--207 (1970).
Halbach, U.: Eilfflu~ der Nahrungsqualit~t und -quantit~t auf die Populations- dynamik des planktischen R~dertieres Brachionus calyciflorus im Labor und
im Freiland. Zoo]. Anz., Suppl. (1971a, im Druck).
Halbaeh, U. : Zum Adaptivwert der zyldomorphen Dornenbildung yon Brachionus calyci]lorus Pallas (P~otatoria). L R~uber-Beute-Beziehung in Kurzzeit-Ver- suchen. Oecologia (Berl.) 6, 267---288 (1971b).
Hutchinson, G. E. : Circular causal systems in ecology. Ann. N.Y. Acad. Sci. ~0, 221--246 (1948).
16"

222 U. Halbaeh und It. J. Burkhardt: Simulation oszillierender Populationen
l~abinovich, J. E.: The appheability of some population growth models to a single- species laboratory population. Ann. entomol. See. Amer. 62,437-442 (1969).
Syn, W. M., Wyman, D. G.: DSL/90 digital simulation language. User's guide. IBM Share Program Library.
Wangersky, P. J., Cunningham, W. J. : On time lags in equations of growth. Prec. nat. Aead. Sei. (Wash.) 42, 699--702 {1956).
Wangersky, P. J., Curmingham, W. J. : Time lag in population models. Cold Spr. Harb. Syrup. quant. Biol. 22, 329--338 (1957).
Dr. Udo Italbaeh Zoologisches Institut der Universit~t D-8000 Miinehen 2, Luisenstr. 14 Deutschland
H. J. Burkhardt Deutsehes Rechenzentrum D-6100 Darmstadt, Rheinstr. 75 Deutschland