Embed Size (px)
Citation preview

Z. Lebensm. Unters.-Forsch. 163, 213--215 (1977)
Zeitschrift fur
Lebensmittel- Untersuchung
und -Forschung @ J. F. Bergmann-Verlag 1977
Uber den enzymatischen Abbau yon Tripolyphosphat und Diphosphat in zerkleinertem Fleisch
VI. Einflull des pH-Wertes auf die Tripolyphosphatase- und Diphosphatase-Aktivitiit im Rindermuskel*
Reiner H a r e m und R u n e Neraal
[nstitut f/Jr Chemie und Physik der Bundesanstalt fiir Fleischforschung, Blaich 4, D-8650 Kulmbach
On the Enzymatic Breakdown of Tripolyphosphate and Diphosphate in Minced Meat VI. Influence of pH on the Tripolyphosphatase and Diphosphatase Activities in Bovine Muscle
Summary. The pH o p t i m u m of the t r ipolyphosphatase
activity of minced bovine muscle post rigor was 5.6. The pH o p t i m u m of the d iphosphatase activity was in the range between pH 6.7 and 6.8. The prolonged in- fluence of pH values a r o u n d 7 caused an addi t ional increase in the d iphosphatase activity.
Zusammenfassung. Das p H - O p t i m u m der Tripoly- phosphatase-Aktivit~it in zerkleiner tem Rindermuske l post rigor lag bei pH 5,6, dasjenige der Diphosphatase- Aktivit~it zwischen p H 6,7 und 6,8. Durch lgngere Ein- wi rkung yon pH-Wer t en um 7 wurde die Diphospha- tase-Aktivit~it zusfitzlich erhSht.
Einleitung
Wie wir in friiheren Untersuchungen gezeigt haben, vergndert sich die Aktivitfit sowohl der Tripolyphosphatase (TPase) als auch der Diphosphatase (Dpase) im Muskelgewebe nach dem Schlachten des Tieres in charakteristischer Weise [1, 2]. Da w~ihrend der Lagerung des Muskels post mortem (p.m.) der pH-Wert des Gewebes von Wer- ten um 7 auf 5,5 absinkt, war die Frage von Interesse, ob diese pH- Abnahme far die Anderung der TPase- und DPase-Aktivit~it 1 ver- antwortlich ist. Aus diesem Grunde wurde der Effekt des pH-Wertes auf die Aktivit~it dieser Enzyme im zerkleinerten Rindermuskel stu- diert. Hierzu wurde der Rindermuskel nach Eintritt des Rigor mor- ris zerkleinert und mit NaOH oder HC1 auf verschiedene pH-Werte zwischen 5 und 8 eingestellt. In diesen Proben wurde der Abbau yon zugesetztem Tripolyphosphat (TP) bzw. Diphosphat (DP) verfolgt.
* Diese Arbeit ist Teil der Dissertation yon R.Neraal (Universitg_t Giel3en, 1975)
Abktirzungen: TP = Tripolyphosphat; DP = Diphosphat; TP- ase=Tripolyphosphatase; DPase=Diphosphatase; ATP=Adeno- sintriphosphat; ATPase=Adenosintriphosphatase; p.m.=post mortem
Da in der Praxis Phosphate in Kombination mit Kochsalz ange- wendet werden, wurden die Enzymaktivit~iten in Gegenwart yon 2% NaCI hestimmt.
Material und Methoden
Den M. Iongissimus dorsi des Rindes dem vier Tage nach dem Schlachten bei + 4 ° C abgeh/ingten TierkFrper entnehmen, yon an- h/ingendem Fett- und Bindegewebe befreien und im Fleischwolf (3 mm-Scheibe) zerkleinern. Proben des zerkleinerten Muskels vor dem Phosphatzusatz unter griindlichem Vermischen mit 0,5 n-HC1 oder 0,5 n-NaOH auf verschiedene pH-Werte zwischen 5 und 8 ein- stellen, dann entionisiertes Wasser zumischen. Gesamtzusatz an Fltissigkeit: 25%. Vorversuche batten ergeben, dab die eingestellten pH-Werte verh/iltnism~fl3ig konstant blieben, dab aber bei h6heren pH-Werten innerhalb der ersten Stunde nach Einstellung eine ge- wisse Abnahme eintrat, wonach der Wert sich nicht mehr/inderte. pH-Einstellung mit NaOH bzw. HC1 und nicht mit Pufferl6sungen, da wegen des starken Pufferungsverm/Sgens des Muskelgewebes so hohe Pufferkonzentration erforderlich w/ire, dab eine Beein- flussung der Enzymaktivit~iten durch die Fremdionen des Puf- fers zu beftirchten ist. - - Obwohl mit dem verwendeten Digital- pH-Meter PHM 52 (Firma Radiometer, Copenhagen) der pH-Wert auf die zweite Dezimale genau gemessen werden konnte, lag der Streuungsbereich kaum unter ± 0,05 pH-Einheiten.
Nach pH-Einstellung den Proben 0,5% TP (NasP3010) bzw. 0,5% DP (Na4P2OT) zusetzen; hierdurch pH-Erh6hung um 0,1 bis 0,6 pH-Einheiten; diese war mit DP h6her als mit TP und im unteren pH-Bereich grSl3er als im h6heren. Die den Abb, i und 2 zugrundelie- genden pH-Werte waren nach Zusatz der Phosphate gemessen wor- den. Messung der TPase- und DPase-Aktivit~it bei 22 ° C nach dem friiher angegebenen Verfahren [3].
Ergebnisse nnd Diskussion
1. pH-Optimum der Tripolyphosphatase-Aktivit&
A bbi l dung 1 zeigt den Einflu/3 des pH-Wer tes auf die TPase-Aktivit~it des zerkleiner ten Rindermuskels post rigor in Gegenwar t von 2% NaC1. Ein ausgepr~igtes p H - O p t i m u m der Enzymaktivi t / i t lag bei 5,6. Dieses Ergebnis war reproduzierbar (Wiederholung mit dem Muskel zweier weiterer Rinder). Die starke Aktivit&its- zunahm¢ mit s inkendem pH - W e r t im Bereich zwi-
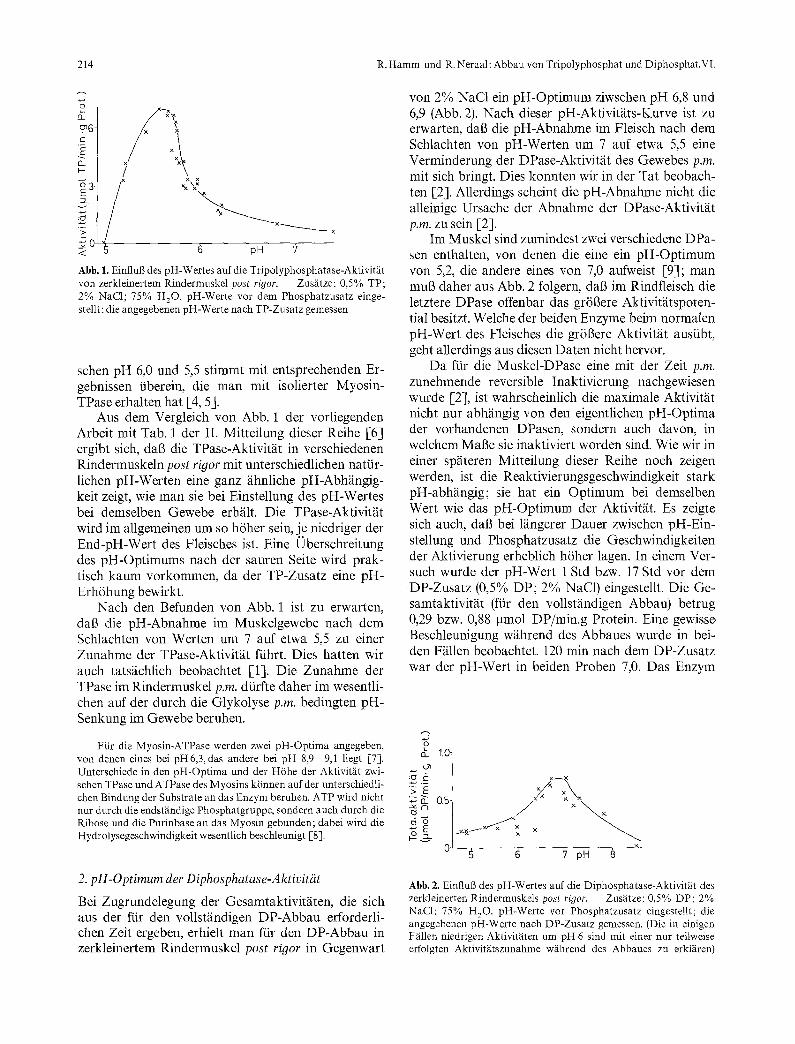
214 R. Hamm und R. Neraal: Abbau von Tripolyphosphat und Diphosphat.VI.
+3 /3_
9~6- e-
o_
5 ._>
X,,xX Xxx
6 pH 7
Abb. 1. EinfluB des pH-Wertes auf die Tripolyphosphatase-Aktivit~it von zerkleinertem Rindermuskel p o s t r i gor . - - Zusfitze: 0,5% TP; 2% NaC1; 75% HzO. pH-Werte vor dem Phosphatzusatz einge- stellt; die angegebenen pH-Werte nach TP-Zusatz gemessen
schen pH 6,0 und 5,5 stimmt mit entsprechenden Er- gebnissen iiberein, die man mit isolierter Myosin- TPase erhalten hat [4, 5].
Aus dem Vergleich von Abb. 1 der vorliegenden Arbeit mit Tab. 1 der II. Mitteilung dieser Reihe [6] ergibt sich, dab die TPase-Aktivit~it in verschiedenen Rindermuskeln post rigor mit unterschiedlichen nattir- lichen pH-Werten eine ganz ~ihnliche pH-Abh~ingig- keit zeigt, wie man sie bei Einstellung des pH-Wertes bei demselben Gewebe erh~ilt. Die TPase-Aktivit~it wird im allgemeinen um so h6her sein, je niedriger der End-pH-Wert des Fleisches ist. Eine Oberschreitung des pH-Optimums nach der sauren Seite wird prak- tisch kaum vorkommen, da der TP-Zusatz eine pH- Erh6hung bewirkt.
Nach den Befunden yon Abb. 1 ist zu erwarten, dab die pH-Abnahme im Muskelgewebe nach dem Schlachten von Werten um 7 auf etwa 5,5 zu einer Zunahme der TPase-Aktivit~it fiihrt. Dies hatten wir auch tats~ichlich beobachtet [1]. Die Zunahme der TPase im Rindermuskel p.m. dtirfte daher im wesentli- chen auf der durch die Glykolyse p.m. bedingten pH- Senkung im Gewebe beruhen.
Fiir die Myosin-ATPase werden zwei pH-Optima angegeben, yon denen eines bei pH6,3, das andere bei pH 8,9--9,1 liegt E7]. Unterschiede in den pH-Optima und der Hbhe der Aktividit zwi- schen TPase und ATPase des Myosins kgnnen auf der unterschiedli- chen Bindung der Substrate an das Enzym beruhen. ATP wird nicht nur durch die endsdindige Phosphatgruppe, sondern auch durch die Ribose und die Pnrinbase an das Myosin gebunden; dabei wird die Hydrolysegeschwindigkeit wesentlich beschleunigt [8].
2. pH-Optimum der Diphosphatase-Aktivitiit
Bei Zugrundelegung der Gesamtaktivit~iten, die sich aus der ftir den vollst~indigen DP-Abbau erforderli- chen Zeit ergeben, erhielt man ftir den DP-Abbau in zerkleinertem Rindermuskel post rigor in Gegenwart
von 2% NaC1 ein pH-Optimum ziwschen pH 6,8 und 6,9 (Abb. 2). Nach dieser pH-Aktivit~its-Kurve ist zu erwarten, dab die pH-Abnahme im Fleisch nach dem Schlachten yon pH-Werten um 7 auf etwa 5,5 eine Verminderung der DPase-Aktivit~it des Gewebes p.m. mit sich bringt. Dies konnten wir in der Tat beobach- ten [2]. Allerdings scheint die pH-Abnahme nicht die alleinige Ursache der Abnahme der DPase-Aktivit~it p.m. zu sein [2].
Im Muskel sind zumindest zwei verschiedene DPa- sen enthalten, von denen die eine ein pH-Optimum von 5,2, die andere eines von 7,0 aufweist [9]; man mul3 daher aus Abb. 2 folgern, dab im Rindfleisch die letztere DPase offenbar das gr6Bere Aktivit~itspoten- tial besitzt. Welche der beiden Enzyme beim normalen pH-Wert des Fleisches die gr6Bere Aktivit~it austibt, geht allerdings aus diesen Daten nicht hervor.
Da ftir die Muskel-DPase eine mit der Zeit p.m. zunehmende reversible Inaktivierung nachgewiesen wurde [2], ist wahrscheinlich die maximale Aktivit/it nicht nur abh~ingig yon den eigentlichen pH-Optima der vorhandenen DPasen, sondern auch davon, in welchem Mage sie inaktiviert worden sind. Wie wir in einer spiiteren Mitteilung dieser Reihe noch zeigen werden, ist die Reaktivierungsgeschwindigkeit stark pH-abh~ingig; sie hat ein Optimum bei demselben Wert wie das pH-Optimum der Aktivit/it. Es zeigte sich auch, dab bei Wngerer Dauer zwischen pH-Ein- stellung und Phosphatzusatz die Geschwindigkeiten der Aktivierung erheblich h6her lagen. In einem Ver- such wurde der pH-Wert 1 Std bzw. 17 Std vor dem DP-Zusatz (0,5% DP; 2% NaC1) eingestellt. Die Ge- samtaktivitiit (ffir den vollst~indigen Abbau) betrug 0,29 bzw. 0,88 gmol DP/min.g Protein. Eine gewisse Beschleunigung w~ihrend des Abbaues wurde in bei- den F~illen beobachtet. 120 rain nach dem DP-Zusatz war der pH-Wert in beiden Proben 7,0. Das Enzym
o
m 1.0
' .~_ 0.5
¢j o
0
× - - x
6 "l pH
Abb. 2. Einflul3 des pH-Wertes auf die Diphosphatase-Aktivit~it des zerkleinerten Rindermuskels p o s t r igor . - - Zus~itze: 0,5% DP; 2% NaC1; 75% H20. pH-Werte vor Phosphatzusatz eingestellt; die angegebenen pH-Werte nach DP-Zusatz gemessen. (Die in einigen FM1en niedrigen Aktivitfiten um pH 6 sind mit einer nur teilweise erfolgten Aktividitszunahme wghrend des Abbaues zu erkl~iren)

R. Hamm und R. Neraal: Abbau yon Tripolyphosphat und Diphosphat. VI. 215
wird also nicht nur durch das Substrat, sondern auch allein durch ejne Erh6hung des pH-Wertes aktiviert. Analoges war auch bei inaktivierter Hefe-DPase beob- achtet worden [10].
Literatur
1. Hamm, R., NeraaI,R.: Z. Lebensm. Unters.-Forsch. IV. Mitt. 163, 126 (1977)
2. Neraal, R., Hamm, R. : Z. Lebensm. Unters.-Forsch. V. Mitt. 163, 208 (1977)
3. Neraal, R., Hamm, R.: Z. Lebensm. Unters.-Forsch. I. Mitt. 163, 14 (1977)
4. Azuma, N., Ikehara, M., Ohtsuka, E., Tonomura, Y.: Biochim. Biophys. Acta 60, 104 (1962)
5. Yasui, T., Fukazawa, T., Takahashi, K., Sakanishi, H., Hashimo- to, Y.: J. Agr. Food Chem. 12, 399 (1964)
6. Neraal,R., Hamm,R.: Z. Lebensm. Unters.-Forsch. II. Mitt. 163~ 18 (1977)
7. Engelhardt, V. A. :Advances in Enzymol. 6, 147 (1946) 8. Tonomura, Y., Imamura, K., Ikehara, M., Uno, H., Harada, F.: J.
Biochem. 61,460 (1967) 9. Nakamura, S., Yamaguchi, M., Morita, J., Yasui, T. : Japan. B uli.
Meat, Meat Products 5, 19 (1969) I0. Rafter, G.W., Witherspoon, B.H.: Life Sci. 11, 995 (1972)





























