Embed Size (px)
Citation preview

Ueber den specifischen Sauerstoffgehalt des Blutes.' Von
Christian Bohr.
In den vorhergehenden Abhandlungen uber das krystallisirte Hilmoglobin haben wir gesehen, dass es, unter iibrigens gleichen Be- dingungen, nicht immer per Gramm die gleiche Menge von Sauerstoff absorbirt. Wir werden in dieser Abhandlung zeigen, dass auch der in den Blutkiirperchen noch eingeschlossene Farbstoff sich in analoger Weise verhalt.
Wenn wir, nachdem wir das Blut mit atmosphilrischer t u f t ge- schiittelf haben, die Menge von Sauerstoff, welche es absorbirt hat, oder wie man es oft nennt, die respiratorische CapacitZit des Blutes messen, und wenn wir zu gleicher Zeit die Menge des Farbstoffes be- stimmen , finden wir , dass das Verhiiltniss zwischen diesen beiden Griissen nicht constant ist, wie man im Allgemeinen annimmt. Man eflahrt im Gegentheil, dass dieses Verhiiltniss, welches wir den speci- fischen Sauerstoffgehalt des Blutes nennen werden, ein sehr ver- iinderlicher ist bei verschiedenen Individuen derselben Art, zu deren Untersuchung die Gelegenheit sich dargeboten hat. Man sieht ausserdem, dass dieser specifische Sauerstoffgehalt bei einem gegebenen Individuum sich durch Einfliisse verschiedener Natur modificiren liisst, und dass endlich das zu gleicher Zeit verschiedenen Gefass- systemen desselben Thieres entnommene Blut in Bezug auf dieses Ver- hiltniss nicht identisch ist. Diese bisher nicht beobachteten Veriinde- rungen, welchen der Farbstoff des Blutes unterworfen ist, sind nicht
Der Redaction mgegangen den 28. Mgrz 1891.

102 CHRISTIAN ROHE:
ohne Bedeutung fur die Lehre von der Respiration, denn, wie wir melir im Detail im zweiten Capitel es zeigen werden, hangt die Spannung der Gase des Blutes nicht dlein von der Rfenge derselben im Blute, sondern auch von dem augenblicklichen Zustande des Farbstoffes ab. Aber be- vor wir diese Frage naher untersuchen werden, werden wir im ersten Capitel diejenigen Erfahriingen mittheileu , auf melchen die eben aus- gesprochene Anschauung fusst.
Erstes Capitel .
D i e M e t h o d en. Um im Blute das Verhaltniss zwischen bei Atmospharendruck absorbirtem Sauerstoff und der Menge des Farb- stoffes zu bestimmen, habe ich in folgender Weise experimentirt:
Dem Thiere , welches der Untersuchung unterworfen, entnimmt man 100ccm Blut, welches man filtrirt, nachdem man es zur Defibri- nation geschiittelt hat. Das filtrirte Blut wird demnachst 20 Minuten hindurch ununterbrochen bei 15 O in einem Ballon geschiittelt, durch welchen man einen stark stromenden Zug atmosphirischer Luft aspirirt, welcher das Blut unter einem partiellen Drucke von ungefahr 150mm mit Sauerstoff sattigt. Darauf entgast man eine Probe des Blutes und analysirt die erhaltene Luft. Bei den spater beschriebenen Gas- analysen ist in ungefahr der Halfte der Falle eine Modification der Methode von Bunsen und in den anderen die Methode von Pe t te r - son angewendet worden. In dem iibrigen Blute ist gleichzeitig der Eisengehalt, die Lichtabsorption und das Residuum festgestellt worden und um die durch die Analysen benothigten Umrechnungen der Ge- wichte in Volumen zu ermbglichen, gleichzeitig die Dichte des Blutes bestimmt worden ; mit Riicksicht auf die angewendeten Methoden Fer- weise ich auf die Abhandlung iiber die Verbindungen zwischen dem Hamoglobin und dem Sauerstoff. Die quantitative Spectralanalyse ist zum Theil mit dem Apparahe von Glan, zum Theil mit dem von Vierordt-Kriiss ausgefuhrt worden. Indem der Zufall mit sich ge- fiihrt, dass hei den beiden Apparaten nicht in der ganz gleichen Region des Spectrums operirt worden ist, sind schon aus diesern Grunde die Resultate, um untereinander vergleichbar zu sein, stets einer Correction bedurftig.
Die Eisenmenge ist nur in gewiseen Fallen zweimnl in jeder Blut- probe bestimmt worden, namlich in einigen Fallen, in welchen die Resultate auffielen; andererseits ist im Allgemeinen eine Reihe Unter- suchungen uber dieselbe Frage an verschiedenen Individuen einer und derselben A r t unternommen worden.

UEBER DEN SPECIFISCHEN SAUERSTOFFGEITALT DES BILJTES. 103
Um die Menge des Farbstoffes festzustellen, hat man fruher theils den Eisengehalt, theils die Lichtabsorption des Blutes als Grundlage benutzt, namlich in der Voraussetzung, dass diese zwei Bestimmungs- weisen, exact ausgefuhrt, identische Resultate geben miissten.
Aber durch unsere Untersuchungen iiber das Hamoglobin in den vorhergehenden Abhandlungen haben wir schon erfahren, dass der Eisengehalt sowie die Lichtabsorption in dem Hiimoglobin nicht con- stant sind; in Folge dessen kann die mit Hiilfe dieser beiden Grossen ausgefuhrte Berechnung der abs oluten Hamoglobinmenge keine sichcre werden, aber, und dieses ist yon noch grbsserer Bedeutung, wir diirfen die Bestimmung des Eisens und die der Lichtabsorption eben- sowenig bei unseren Untersuchungen iiber den Farbstoff in den Blut- kijrperchen wie bei unscren Untersuchungen iiber die verschiedenen Arten des Krystallhamoglobins als gleichwerthige Methoden in An- wendung bringen; hier wie dort diirfen die beiden Methoden hiich- stens als sich gegenseitig ergiinzende Anwendung finden. Das Ver- hiiltniss zwischen dem Eisengehalt und der Lichtabsorption des Blu tes i s t i n Wirkl ichkei t kein constantes. Dieses Ver- hiiltniss wird dargestellt durch cc,, in der Gleichung Fe = u,,E, wo E den Lichtabsorptionscoefficienten bezeichnet und Fe das in 100 wm Blut enthaltene Eisen.' Folglich besteht auch kein einfaches Verhaltniss zwischen den Sauerstoffmengen, welche im Blute bezw. 113 Eisen und einer Einheit absorbirten Lichtes entsprechen; letzterer Werth ist aus- gedriicbt durch ao,, welches hervorgeht aus der Gleichung: 0 = ao,E, wo E den Absorptionscoefficienten bezeichnet und 0 die 100ccm ab- sorbirte Sauerstofhenge.
Weil also die Verhiiltnisse zwischen dem Eisen und dem Sauer- doff, wie auch zwischen dem Sauerstoff und dem absorbirten Lichte beide im Blute veranderlich sind, und indem ferner, wie wir in einer vorhergehenden Abhandlung erfahren und auch splter nooh ersehen werden , das aus verschiedenen Blutproben erhaltene Hamoglobin in diesen Verhaltnisscn auch nicht constant ist, muss die Vorstel lung aufgegeben werden, dass der Farbstoff des Blutes sich i n Bezug auf die Menge von absorbirtem Sauerstoff constant verhal t , wed diese Vorstellung den Resultaten aller Untersuchungen widerspricht.
Soviel mir bekannt, findet sich in der Litteratur keine Unter- suchungsreihe, welche den Zweck hatte, die hier erorterte Rage zu erhellen, und somit ist ein directer Vergleich zwischen den Resultaten,
Vgl. Ueber die Verbindmgen a h Hamoglobins mit Sauerstoff. S. 86.

104 CHRISTIAN BOHR:
welche ich erhalten und den von anderen Verfassern veroffentlichten Arbeiten iiber das Hamoglobin nicht moglich.
Um nun zu wissen, nach welchem Maassstabe wir den specifischen Sauerstoffgehalt des Blutes oder die bei Atmospharendruck von 1 8 des Farbstoffes absorbirte Sauerstoffmenge zu bemessen haben , wird es nothwendig werden, jede der beiden Methoden (Eisenbestimmung und Spectralanalyse) fur sich zum Gegenstand der Untersuchung zu machen.
Eisenbestimmung. Wir werden sehen, dass das Verhaltniss zwischen der von dem Blute bei Atmospharendruck und 1 5 O absorbirtem Sauerstoffmenge und der Eisenmenge, welche es gleichzeitig enthalt, bei verschiedenen Individuen ein sehr veranderliches ist, sowie unter wechselnden iiusseren Umstiinden auch bei demselben hdividuum. Jeder kann sich mit Leichtigkeit davon uberzeugen, dass die Dosirung des Eisens und des Sauerstoffes des Blutes sich bei einiger Uebung mit grosser Genauigkeit ausfuhren lks t , die bedeutenden Variationen, welche das Verhaltniss zwischen dem Sauerstoffe und dem Eisen (Sauer- stoff per Gramm Eisen) darbietet, sind nicht durch Pehler der Analyse erklarbar. Indem wir uns aber dieses erklaren wollen, miissen wir zu allererst die Frage aufwerfen, ob alles in dem Blute enthaltene Eisen in dem Farbstoffe eingeschlossen ist, falls es sich anders verhielt und sich in dem Blute anderweitig eine Menge von Eisen v o r h d e , die geniigend ware, um einen wesentlichen Einfluss auf die Grosse des Ver- haltnisses zwischen Sauerstoff und Eisen zu haben, wurde die Berech- nung dieses Verhlltnisses nicht die geringste Bedeutung haben, indem vielleicht gar keine Art von Verhiiltniss zwischen einem Theile des Eisens und dem in lose Verbindung mit dem Blute getretenen Sauer- stoff bestehen wiirde.
dber iiltere Verfasser haben im Plasma’ des Blutes nur so kleine Mengen von Eisen gefunden, dass dieselben von gar keinem Einfluss auf unsere Untersuchungen sein konnen; deswegen diirfen wir es aber nicht als iiberhaupt unmoglich ansehen, dass das Plasma einzelne Male Eisenverbindungen in vie1 hoherer Proportion einschliessen konne, ob- gleich solchea bisher niemals nachgewiesen worden ist. Indem nun unsere Untersuchungen zum grossten Theil keine vereinzelt dastehende Beispiele darbieten, sondern aus langen mit der grossten Regelmbig- keit aufeinander folgenden Reihen bestehen, muss dieses, wie es
1 In dem Stroma der KSrperchen des Blutes hat sich kein vorhandenes Eisen nachweisen lassen. Schliisse ea iibrigens Spuren von Eisen ein, wiirde solches ohne Einfluss auf unsere Untersuchungen sein, weil die im Rlute ent- haltene Menge von Stroma eine unbedeutende iet.

UEBER DEN SPECIFISCHEN SAUERSTOFFGEHALT DES BLUTES. 105
scheint, schon genugen, um den Gedanken auszuschliessen, dass unsere Resultate Folgen sein sollten eines zufalligen Vorhandenseins in dem Plasma von bisher unbekannten Eisenverbindungen. Was aber die Rage entscheidet, ist das, was aus den sptter dargestellten Erfahrungen hervorgeht, namlich, dass die fur verschiedene Blutproben nachgewiesenen Unterschiede in dem Verhaltnisse des Sauer- stoffes zum Eisen sich i n dem diesen Proben entnommenen kr y s t a1 lis i r t e n Ham o g 1 o b in w ie d e r f i n d e n.
Dieses Resultates wegen mussen wir, was auch schon von vorn- herein als richtig erscheinen durfte, fur das Blut einen Wechsel in dem Verhaltnisse zwischen dem Sauerstoff und dem Eisen diesel be Bedeutung beilegen, welche wir schon fruher einem analogen Wechsel des dem Blute in Krystallen entnommenen Hamoglobins beigelegt haben. In Bezug auf diese letztere Substanz haben wir aber gefunden,' dass sie eine Mischung mehrerer Hamoglobinarten sei, der Hamoglobine a, /3, y , 6 oder anderer analoger Arten, die, bei einem gegebenen Drucke, im Verhaltnisse zu ihrem Eisengehalt verschiedene Yengen von Sauerstoff absorbiren. Eine Steigerung (oder eine Verringerung) der Sauerstohenge per Gramm Eisen musste demnach, ebensowohl in dem Blute als in der Mischung von Haoglobinen, als Zeichen eines reichlicheren Gehaltes von verhaltnissmissig mehr (oder weniger) Sauer- stoff absorbirenden Hamoglobinen betrachtet werden. Die Bedeutung, welche unsere Untersuchungen hierdurch fur die Theorie der Respiration erhalten, wird in dem zweiten Capitel dieser Abhandlung dargelegt werden.
[Es ist hier der Ort auf eine Lucke in meinen Untersuohungen fiber die versohiedenen Hamoglobine die Aufmerksamkeit zu lenken. Indem ihr Eisengehalt ein verschiedener sein kann, wurde der Fall eintreten konnen , dass zwei Mischungen verschiedener Hiimoglobine die gleiche Menge von Sauerstoff per Gramm Residuum, eine verschie- dene Menge jedoch per Gramm Eisen absorbiren. Es wiirde hier sweifelhaft bleiben, ob zwei derartige Hamoglobine in Bezug auf die Spannung, welche sie einer bestimmten Menge von absorbirtem Sauer- stoff mittheilen, identisch sind (in Folge Residuumsbestimnung) oder aber verschieden (in Folge Eisenbestimmung). Dass letztere Alter- native die rechte sei, scheint mir wahrscheinlich, weil der gefirbte Kern, welcher den Sauerstoff bindet, zu gleicher Zeit das Eisen ent- h a t und weil die Menge des Residuums theilweise von dem ungefirbten Theil des Hamoglobins abhingig ist, welcher keinen Theil an der Bin-
Vgl. Bohr, Ueber die Verbindungen des Hamoglobins wit Sawstof.

106 CHRISTIAN BOHR:
dung des Sauerstoffes hat. Ich habe diese Frage noch nicht experi- mentell untersucht. Die Untersuchung ware yielleicht in der Weise vorzunehmen, dass man das Verhaltniss zwischen gebundenem Sauer- stoff und Eisen vor und nach der theilweisen PBllung des ungefirbten Theiles des Hamoglobinmoleciils bestimmt, es bleibt aber fraglich, ob in Folge mijglicher gleichzeitiger Veriinderungen des gefiirrbten Theiles dieser Weg zu einem sicheren Resultate fuhren wurde.
Fur die Untersuchungen, mit denen wir uns in dieser Abhand- lung beschiftigen, hat diese Frage nur eine geringere praktische Be- deutung, da hier der Sauerstoff, welcher in den Hlnioglobinen, die wir zu vergleichen haben werden, per Gramm des Eesiduums odes des Eisens gebunden ist , in demselben Sinne wechselt , obgleich dieser Wechsel in einem Falle rerschiedenen Werth hat.]
Spectralanalyse. Die Menge Sauerstoffs, welche im Blute einer Einheit absorbirten Lichtes entspricht, und welche durch aoz (ver- gleiche oben) ausgedruckt ist, ist ganz so veranderlich als die Nenge, welche per Gramm Eisen gebunden ist. Aber die Veranderungen dieser beiden' Werthe folgen nicht demselben Gesetze. Dieses stimmt mit unseren Untersuchungen uber die verschiedenen krjstallisirten Hamo- globine, denn indem mir in Bezug auf die beiden Hamoglobine y und /? einen exacten Vergleich haben anstellen khmen, haben wir erfahren, dass der Sauerstoff per Einheit absorbirten Lichtes in den verschie- denen Hamoglobinen keineswegs proportionell ist mit dem per Gramm Eisen gebundenen Sauerstoff.' Da nun eine wechselnde Menge Sauer- stoff per Gramm Eisen in dem Blute, wie wir eben gesehen haben, aus der wechsdnden Menge der verschiedenen Hamoglobine, welche es ent- hd t , hervorgeht, ist es hiermit in guter Uebereinstimmung, drsss a,, und der Sauerstoff per Gramm Eisen in dem Blute ebensowenig wie im Krystallhlmoglobin proportionell sind. Im Gegentheil hat man zu erwarten, dass man in verschiedenen Blutproben zwischen dem Eisen und der Lichtabsorption dieselbe Relation vorfindet, die wir in den Hamo- globinen nachgewiesen haben , welche verschiedene Mengen Sauerstoffs binden wachst, wenn der Sauerstoff per Gramm Eisen abnimmt).a Dies geht auch, was die Hauptpunkte angeht, aus unseren Unter- suchungen hervor, wie spliter gezeigt werden soll. Jedoch die werth- volle E r g h u n g , welche dieser Umstand uns in Bezug auf die Be- deutung unserer Eisenbestimmungen an die Hand geben, bietet die grosse Lucke dar, dass die Regelmlssigkeit, welche letztere auszeichnet,
Vgl. Ueber die Verbindungem 02s Hamoglobins mit Sazcerstof. S. 81 ff. Eb&. S. 88.

UEBER DEN SPECIFISCHEN SAUERSTOFFGCEHAhT DES BLUTES. 107
bei den Spectraluntersuchungen fehlt; man sieht hier viele bedeutende Abweichungen von den allgemeinen Regeln, die wir aufzustellen im Stande sind, was zweifellos in Verbindung steht mit den Unregel- missigkeiten in Bezug auf Lichtabsorption, welche yon Zeit zu Zeit auch in den Versuchen iiber das Hamoglobin vorkommen.'
Die Ursachen dieser Abweichungen auszufinden ist um so schwie- riger, weil wir im Gegensatze zu dem, was wir in Bezug auf per Gramm Eisen absorbirtem Sauerstoff gefunden haben, bei der Lichtabsorption die Unterschiede an den verschiedenen Blutproben nicht miederfinden, in dem aus denselben dargestellten Hamoglobin. Blutproben, welche in Bezug auf Sauerstoff und Eisen die gleiche Lichtabsorption ergeben haben, kiinnen Hamoglobine liefern, welche in dieser Beziehung sehr bedeutende Unterschiede darbieten und umgekehrt.
Vielleicht mogen einige der Unregelmassigkeiten in der Licht- absorption in mit den Experimenten verbundenen technischen Schwierig- keiten ihren Grund haben, darauf werden wir gleich zuriickkommen. Eine derartige Erklarung ist aber oftmals sehr unwahrscheinlich, bei- spielsweise in dem besonderen, sehr genau beobachteten Falle, wo das arterielle Blut und allein dieses durch eine Vergiftung mit Cocain in seiner Lichtabsorption in einer Weise modificirt ist, welche ich bei meinen Untersuchungen iiber das Hamoglobin nicht gefunden habe. Thatsachen dieser Art werden uns iibrigens zur Geniige daran erinnern, dass die Untersuchungen, welche wir hier in einer Reihe von Abhandlungen iiber die Eigenschaften der verschiedenen Hamoglobine veroffentlichen, sehr unvollstiindig sind und dass mehr wie ein bedeutsamer Factor zaeifellos unserer Aufmerksamkeit entgangen jst.
In Betreff der oben erwahnten technischen Schwierigkeiten habe ich weniger vor Augen die einfache Bestimmung der Lichtabsorption, welche sich immer geniigend exact wiirde ausfiihren lassen, als die von Torupa gemachte Beobachtung, dass die Hinzugabe von wenig doppeltkohlen- saurem Natron zu einer wasserigen Auflosung von Hamoglobin das Spectrum derselben in dem Sinne modificirt, dass der Punkt der stirksten Absosption ein wenig verschoben ist. Diese Thatsache, welche Torup durch eine Reihe ron genauen Messungen mittels des Photometers von Glan gefunden hat, ist von fundamentaler Bedeutung beziiglich der praktischen Anwendung der Spectralanalyse bei Blutuntersuchungen. Es geht aus derselben in der That herror, davs eine unbedeutende Ver- hnderung des Hamoglobins, welche gar keinen Einfluss hat weder auf
Vgl. H-oglobin Bbdets Kdsgrebinding (Die Xohlemliurebhdung d. Blutes). Copenh. 1866.
a. a. 0. S. 89.

108 CHRISTIAN BOHR:
das Moleculargewicht , noch auf die Menge absorbirten Sauerstoffes, uns in Bezug auf die Lichtabsorption einen ganz anderen Werth an die Hand geben kann, und dieses nicht weil dieselbe in ihrem all- gemeinen Charakter modificirt ist, sondern weil das Absorptionsspectrum ein wenig verschoben worden ist. Ich habe diesem schwierigen Um- stande entgegenzutreten venucht, indem ich immer rnit einer 1/20 pro- centigen Losung von doppeltkohlensaurem Natron gearbeitet habe. Es liegt aber in der Natur der Sache, dass ich durch dieses Mittel nicht den Einfluss habe heben konnen, welchen die vielleicht sehr wech- selnde Menge der in den verschiedenen Blutproben en thaltenen alka- lischen Salze auf das Absorptionsspectrum auszuuben im Stande sind ; es offnet sich hier, scheint mir, ein weites Feld fur Abweichungen in der Lichtabsorption, welche das Aussehen haben, als waren sie die Folge von bedentenden Veranderungen des Hamoglobins, wahrend sie in der That aus Veranderungen hervorgehen, welche klein und fur unsere Zwecke bedeutungslos sind. Man muss immer der Moglichkeit der- artiger Abweichungen gewartig sein, wenn man mit dem Spectral- apparate arbeitet; Torup a macht beispielsweise auch die Bemerkung, man diirfe bei Experimenten mit Blut , welches rnit doppeltkohlen- saurem Natron ,versetzt ist, sich nicht solchen Lichtabsorptionscon- stanten bedienen, die mit Hiilfe von wkserigen Losungen von Hamo- globin zu Wege gebracht sind, wie man haufig thut.
[Eine Beobachtung, welche wir wahrend des Verlaufes dieser Unter- suchungen gemacht haben, scheint anzudeuten, dass die Zeit, welche zwischen der zum Zweck der Spectralanalyse vorgenommenen starken Verdunnung des Blutes und der Spectralanalyse selber verlauft, auch nicht immer ohne Einfluss auf das Absorptionsspectrum sei; iiber diesen Punkt weiss ich aber nichts sicheres zu sagen, indem ich bisher noch keine Gelegenheit gehabt habe, dariiber eine systematische Untersuchung anzustellen.]
...
Bus dem oben iiber die Eisen- und Spectralbestimmung Ent- wickelten geht hervor, dass wir als Maass fur den specifischen Saner- stoffgehalt des Blutes das Verhiiltniss zwischen dem Sauentoff und dem Eisen in demselben
Vgl. Ueber die Capitel, Tabelle S. 93.
a. a. 0. 6. 48.
zu benutzen haben, wiihrend die Spectralanalyse
Verbin.dungen des Hamogbbina rnit Sawstoff. Zweites
I

UEBER DEN SI'ECIFISCHEN SAUERSTOFFGEHALT DES BLUTES. 109
vorliiufig nur als eine die Eisenbestimmung supplirende Methode sich benutzen 1asSt.
Wenn wir deshalb im Folgenden von specifischem Sauer- s toffgehal te sprechen, wird h ie runter der per Gramm Eisen nn te r den iifter genannten Bedingungen (15O und 150"" Sauer- stoffdruck) aufgenommene Sauerstoff verstanden.
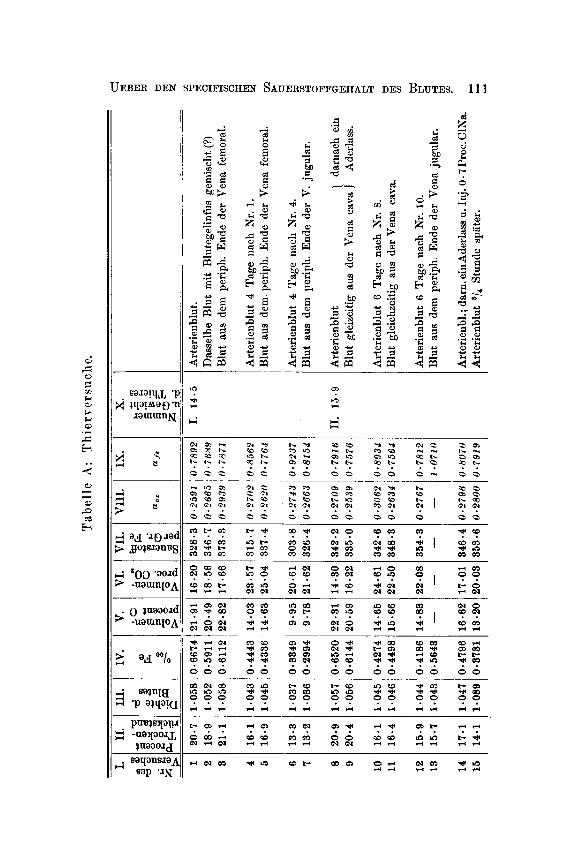
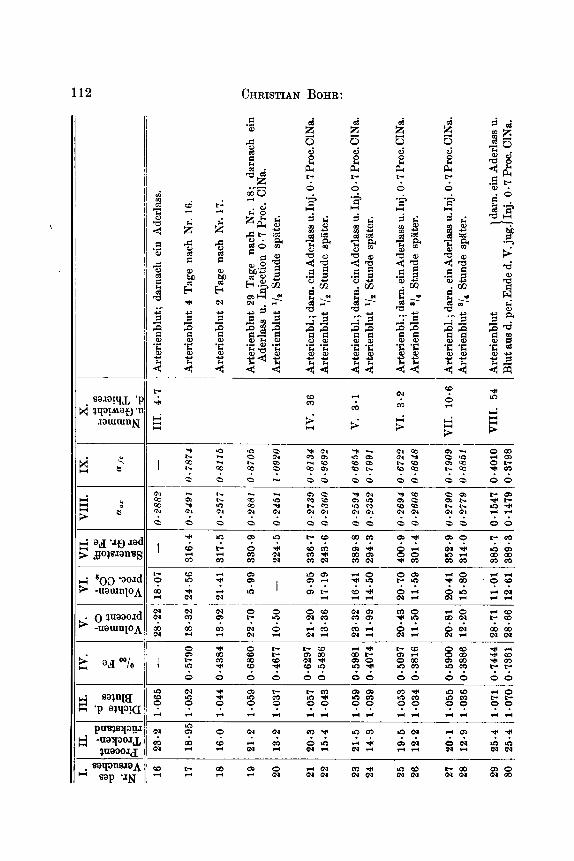
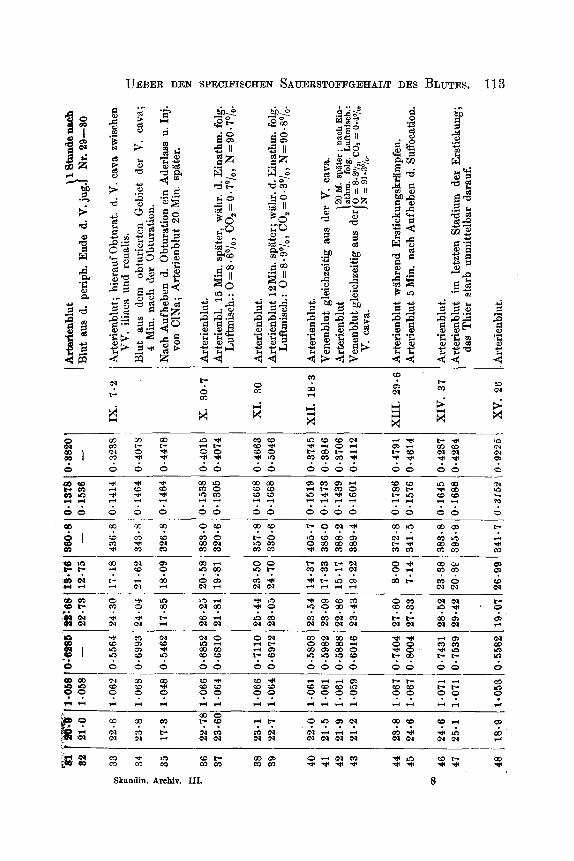
Sammtliche tiber die hier behandelten Fragen angestellten Ver- suche habe ich in untenstehender Generaltabelle aufgefiihrt.
Dieselbe zerfillt in zwei Abtheilungen A und B. A enthalt die an lebenden Organismen angestellten Versuche, B die Versuche in vitro. In der Bbtheilung A finden sich fiir jede einzelne Blutprobe Rubriken fur das Gewicht des Thieres, das Residuum (Gewichtsprocent), Gramm Eisen in 1000 ccm Blut, Volumenprocent Sauerstoff und Kohlen- siiure bei Oo und 760 mm gemessen, Cubikcentimeter Sauerstoff per 1 g
Fe (specifischer Sauerstoffgehalt), sowie fur die aus den Spectralunter- suchungen hergeleiteten und aus den Gleichungen 0 = a,E und Fe = oc,,E berechneten Grossen cc, und ufe, indem E den Exstinctions- coefficienten, 0 und Fe bezw. Cubikcentimeter Sauerstoff und Gramm Eisen in 100 eem Blut bezeichnen. Da einige der Spectraluntersuchungen mit Glsn's Apparate in der Spectralregion: il = 544, andere mit Vierordt-Kriiss' Apparat in der Spectralregion il = 545 ausgefiihrt worden sind, und da die Versuche jeder dieser Gruppen nur unter sich zu vergleichen sind, sind die mit Glan's Apparat gefundenen .Irshlen mit Cursivschrift gedruckt, die ubrigen mit den gewiihnlichen m e n der Tabelle.
Bei jeder einzelnen Blutprobe findet man angegeben, wie dieselbe zu Wege gebracht worden ist; die zu gleichem Versuche gehorenden Proben sind in einer Gruppe fur sich gestellt und von den iibrigen Gruppen durch einen Zwischenraum geschieden. Die Versuchsthiere, welche stets Hunde gewesen , sind mit romischen Zahlen bezeichnet. Wo in der Tabelle ,,Arterienblut" und ,,Venenblut" ohne nahere Be- stimmung angefiihrt worden ist, ist es Blut von normalen Individuen; wo Proben von Arterienblut und Venenblut unmittelbar in derselben Gruppe auf einander folgen, sind sie soweit wie moglich gleichzeitig genommen. Das ,,Venenblut aus der Vena cava" ist, wo anders nicht ausdriicklich bemerkt, aus der untersten Yartie derselben mittels eines durch die V. femoralis eingefiihrten gewohnlichen elastischen Katheters genommen worden, wiihrend der Probenahme ist der Blutstrom in der Vena cava nicht unterbrochen gewesen.
Die Spermng der Vena cava oder des Arcus aortae geschah mittela ekes Gummiballons, der an's Ende eines diinnen metallenen Katheters

110 CHRISTIAN BOHR:
befestigt, in die genannten Gefisse bezw. durch die Vena und Arteria femoralis eingefuhrt worden ist. Der Gummiballon ist darauf durch das Katheter mittels einer Spritze soweit mit Wasser gefullt worden: dass derselbe das Lumen des Gefiisses sprrrte.
W o angefuhrt ist, dass ein Aderlass mit nachfolgender Injection von 0.7 Proc. Cubikceritimeter ClNa vorgenomiiien worden ist, bedeutet dieses, dass durch eine Vene ein dem verlorenen Quantum Blut ent- sprechendes bis auf Korpertemperatur erwarmtes Volumen der ge- nannten Flussigkeit injicirt worden ist.
Die Generaltalxlle hat den Zweck, dem Leser einen vollstlndigen Einblick in die Versuchsdetaile zu geben; die Uebersicht, welche die- selbe giebt, ist jedoch keine gute, indem die Versuche dem Zeitpunkte ihrer Ausfuhrung und nicht ihrer Art nach gruppirt worden sind. Es wird deshalb der Tabelle eine Reihe von Abschnitten folgen, in denen die Versuche so ubersichtlioh wie miiglich zusammengestellt und so sich sammeln, wie sie natiirlich zusammengehoren.
In der Abtheilung B der Tabelle finden sich die Versuche mit Verdunnungen des Blutes und Zusatzen von Giftstoffen, alles ausser- ha1 b des Organismus; gleichzeitig findet man hier die Untersuchungen uber Hamoglobine, die aus bestimmten von der Abtheilung A der Tabelle herriihrenden Blutproben dargestellt sind. Die Darstellungsweise des Hamoglobins ist die in einer fruheren Abhandlung besprochene. In der Abtheilung B ist an mehreren Orten der Eisengehalt verschie- dener Fliissigkeiten dazu benutzt, die enthaltene Hamoglobinmenge aus- zurechnen, wenn die procentische Eisenmenge letztgenannten Stoffes = 0.37 Procent gesetzt wird. Es kann dieses bei diesen Versuchen kein Nissverstandniss mit sich fuhren und giebt eine mehr gewohnheits- mZissige Ausdrucksweise.
Die Verbindungen des Ha.moglobins nzit Sawrstoff, zweites Capitel.
5 2 E E iisauemtoff ,-j w ww ww PP wu- ww 01P
03P w w03 010 u-w u-4 w-lw 00 0 00 00 00 55 ooc
W '? perGr. Fe w03 I$ wy 0110 .. ..
u 11 Nummer u.Gewiclit 8
;n lid. Thieres
WN NN NN NN NN N + c c ,-
NN cI h3 to - c"c" ?? COY t': c"? W 7 ? ? W
.. .. .. .. .. - 00 00 44 ww g8 gg %8 % 8 P 8
O -I ow w-l -01 PW NC
PP We NOI WCn PW N N 0 CD Cn
el- cc el- cc cc I- c L c c
or a01 PW ww w4 4 w P N OI
Nr.des ,-, Versuches' Procent
Trocken-
Dichte d. y
N&,~~n,j
Blutes
u.Gewicht ? Id. Thieres dl
CC WN cN Cc c NNI- ps': c"? yo p? 4- .. 01 - * 00
' A 6 b 3 wcm zz WP or gE! g5 .zg
woo 1-01 00 wa, PW LO w c I-
t 0 f F ' "c" tpfs I-? pw wm WW WW WP NW COW N W W W
.. w4 ow Pw ww a4 Cn w -a P
Volumen- 4 proc. CO, F
Sauerstoff 2 perGr.Fe F
“di
32
u,
33
F g 34
P
5 a 35
E
Y
l-4
36
37
38
39
40
41
42
43
44
45
00
46
47
48
‘Mm
21
.o
22-6
23.8
17.3
22 * 7
1 23
.6(
23.1
22
.7
22.0
21
.5
21.9
21
.2
23.8
24
-6
24-6
25
-1
18.9
1.05
8 1 - 05
8
1.06
2
1.06
8
1.04
8
1.06
6 1.
064
1.06
6 1
so64
1.06
1 1.
061
1-06
1 1-
059
1-06
7 1.
067
1.07
1 1-
071
1.05
3
0:68
85
-
0 * 5
564
0.69
93
0 * 54
62
0.68
52
0 * 68
10
0.71
10
0.69
72
0.58
03
0 * 59
82
0.58
88
0.60
16
0 * 74
04
0 * 80
04
0.74
31
0.75
39
0 * 5
582
%8:
68
22.7
3
24.3
0
24-0
4
17.8
5
26 *
25
21.8
1
25 - 44
23
* 05
23.5
4 23
.09
22.8
6 23
.43
27.6
0 27
.33
28.5
2 29
.42
19.0
7
18.7
6 12
.75
17.18
21.6
2
18.0
9
20.5
8 19
.81
23 * 5
0 24
.70
14.3
7 17
-33
15 * 1
7 19
.22
8.00
7-
14
23-3
8 20
.3fl
26-9
9
360.
8 -
436.
8
343.
R
326
* 8
383 *
0
320.
6
337
* 8
330.
6
405-
7 38
6.0
388-
2 38
9.4
372-
8 34
1.5
383 *
8
395
* 9
341 *
7
0-13
76
0.15
36
0.14
14
0.14
64
0 * 14
64
0.15
38
0.13
05
0.16
68
0.16
68
0.15
19
0.14
73
0.14
39
0*1
60
1
0.17
86
0.15
76
0.16
45
0.16
88
0.31
52
0 * 3
820
-
0 * 32
38
0.40
75
0.44
78
0.40
15
0.40
74
0 * 46
63
0 - 50
46
0.37
45
0-38
16
0.37
06
0.41
12
0.47
91
0.46
14
0.42
87
0-42
64
0 - 92
25
IX.
7.2
x. 3
0-7
XI.
30
XII
. 18
.3
XII
I. 2
9.6
XIV
. 37
Xv.
26
Blu
t au
s d.
per
iph.
End
e d.
V.
Art
denb
lu t
Arte
rienb
lut ;
hie
rauf
Obt
urat
. d.
V. e
ava
zwis
chen
Blu
t au
s dc
rn
obtii
rierte
n G
ebio
t de
r V
. ca
w;
Nac
h A
ufhe
ben
d. O
btur
atio
n ei
n A
derl
ass
u. I
nj.
a g m
U
VV
. ili
aca
und
rena
lis.
4 M
in.
nach
dcr
Obt
urat
ion.
von
ClN
a; A
rter
ienb
lut
20 M
in.
spat
er.
!2 A
rter
ienb
lut.
% x A
rterie
nblu
t. M
Arte
rienb
l. 15
Min
. spa
ter,
wah
r. d
. Ein
athm
. fo
lg.
Luftm
isch
. : 0 =
8 - 6O
/, , G
O, =
0.7O
/, , N
= 90
. 7O/,
.
b2 !4 M
Art
erie
nblu
t 12
Min
. spa
ter;
wah
r. d.
Ein
athm
. fol
g.
Luftm
isch
. : 0 =
8. g
o/, , c
o, =
0.3O
/, , N =
90.8
O/,.
Art
erie
nblu
t. V
enen
blut
gle
ichz
eitig
aus
der
V.
cava
.
Ven
enbl
ut g
leic
lizei
tig a
us d
er o
s.
30/,,
, cq
= o
.q,,
Q
R E
V. c
ava.
(N
91.3
°/0.
k4 0
lnth
m.
folg
. Lu
ftmis
cb.:
4 A
rter
ienb
lut
20 M.
spst
er ; n
nch
Ein
-
5 A
rter
ienb
lut
wah
rend
Ers
ticku
ngsk
ram
pfen
. U
A
rter
ienb
lut
5 M
iu.
nach
Auf
hebe
n d.
Suf
foca
tion.
Art
erie
nblu
t. A
rter
ienb
lut
iin l
etzt
en S
tadi
um d
er E
rstic
kung
;
W
F
das
Thi
er s
tarb
unm
ittel
bar
dara
uf.
P
Arte
rienb
lut.
=
I. P m
a
mc
&) -
-
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
II.
,T
ig
: z+
jMof
2 o<
4 g
:E -
-
24.5
24
.5
24.6
24
.6
23.6
22.2
21.6
21
.6
21
-5
22.9
22
.2
21.3
20
.7
19.9
1
9-7
23.0
24
.5
-
-
111.
4,
88
g
e
ZcQ
-
-
1 a0
69
1.06
6
1.07
0 1
-069
1.06
7
1-06
3
1.06
1 1.
060
1 -05
9 1.
061
1 -06
3
1.06
0 1.
058
1.05
5 1.
055
1 so6
4 1.
068
-
IV. z 0
0-
0 64
77
0.69
30
0.7808
0.77
48
0.57
73
0.65
76
0.70
24
0.68
81
0.67
99
0.66
96
0.69
91
0.64
66
0.63
56
0.63
08
0.73
43
0.81
61
0 * 6
994
=
V.
QO
a’;i
218 -
28-1
4 28
.16
27.9
1 27
.75
26-6
5
24.7
4
24.0
1 23
- 60
23.6
3 24
.53
24 * 6
5
24-1
3 22
-17
20.6
8 20
.14
26.8
28
.13
-
-
VI.
a0
Q& +
g -
16.6
8 10
-23
19
-20
14
.00
8.71
7-5
1
26.7
9 1
9-0
1
20 * 1
0 11
.78
8.81
14.4
4 6.
50
22 * 6
8 14
.67
11.5
2 5.
34
=
VII
. %
G
-
434.
f 40
6.4
357.
5 35
8.1
461 *
1
37
6-1
341.
9 3
43
-1
347.
5 36
6.4
352
* 6
373
* 1
34
8 * 7
327.
8 27
4 * 3
328
- 6 40
2 * 2
=
VII
I.
a ox
0 * 13
82
0.14
91
0.13
50
0.13
33
0.19
37
0.21
25
0.14
91
0.16
70
0.15
96
0.17
26
0.16
79
0.17
96
0-17
81
9.27
30
9 - 28
26
1.27
13
1.27
34
-
IX.
a /C
o.m
a 0.
3668
0.37
76
0.37
23
0-41
95
0.56
48
0.43
60
0.48
70
0.45
94
0.47
09
0.47
61
0.48
12
0.51
06
1 * 8
327
I a03
0
1.82
59
1.67
97
Art
erie
nblu
t. B
lut
glei
chze
itig
aus
der
V.
cava
.
Art
erie
nblu
t )~
ura
re.
Blu
t gl
eicl
izei
tig a
us d
er V
. ca
va
Art
erie
nblu
t; da
rn.
Obt
urat
ion
zuer
st d
er
Blu
t 4 1
1. sp
lt. a
. d. o
btur
.Geb
iet d
. V. c
ava\
Blu
t gle
ichz
. a. d
. V. c
ava;
hie
rauf
Obt
ur. d
. Aor
ta
2 B
lut a
us d
. V. c
ava
3 N
in. n
acli
Obt
urat
. d. A
orta
$
Blu
t aus
d.V
. cav
a 30
Min
. nac
liob
tura
t. d.
Aor
ta 0
Art
erie
nblu
t
Art
erie
nblu
t 2
Min
. na
ch E
nde
der
Obt
urat
ion
0
m E A
orta
und
dan
n de
r V
. cav
a \C
urar
e. 2 P 4 W
0
M i
Art
erie
nblu
t: hi
erau
f O
btur
atio
n de
r A
orta
. A
rter
ienb
lut w
iihre
nd d
. Obt
nrat
.; 1
St. n
acli
Nr.
60.
Art
erie
nblu
t; hi
erau
f O
btur
atio
n d.
Aor
ta.
Art
erie
nblu
t wah
rend
d. O
btur
at.;
1 St
. nac
h N
r. 62
.
Arte
rienb
lut.
Blu
t au
s de
r V. p
orta
, na
chde
m
dies
e ku
rze
Zeit
unte
rbun
den
war
.
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
",
81
82
23
-4
21.8
22.1
22.0
24.7
2
4-1
24
.6
-
22.1
21
.7
21.5
21.8
20.3
21.9
21
.6
21.6
21.3
1.06
6
1.06
2
1.06
3
1.06
3
1.06
8 1.
068
1.06
9
1 a0
68
1 -0
62
1.06
2 1
-06
1
1 *0
63
1.05
8
1 -06
0 1.
059
1.06
0
1.06
0
0.72
71
0-63
08
0.59
94
0.61
13
0-75
28
0.71
24
0.77
83
-
0 * 62
26
0 - 640
4 0
- 6041
0.64
92
0.56
41
0 * 4
866
0: 54
14
0 * 42
00
0 * 5
427
21-2
2
23.9
7
24.3
9
24.3
5
28-0
8 27
- 30
28 - 12
28 * 8
4
23 * 5
9 23
- 43
23 * 1
5
23.5
3
20 * 9
4
22 * 7
5 22
.71
23.0
7
23.1
6
14 * 3
2
24.5
2
25.3
0
-
13.8
2 17
.14
11 * 5
4
6.58
24-4
2 23
* 84
21
.73
16-8
3
12.8
0
19.0
3 12
.86
19.8
1
14.2
9
374.
3
379
* 9
407
-0
398.
4
373-
0 38
3-2
361.
3
-
379.
0 36
5 * 9
383
* 3
362
* 4
371
-2
467.
5 41
9 * 6
549.
4
426
- 8
1.18
10
)a12
65
)*13
79
b.13
66
)a13
74
)- 13
65
1.13
39
1 * 1
413
1.15
66:
1.16
50
1.12
82
1.15
60
1.12
52
-
-
-
-
1.49
96
1.33
29
) * 3
389
1.34
30
1.36
83
1-35
61
1.37
06
-
1.41
16
) * 4
510
) * 3
346
)a43
05
1.33
74
-
-
-
-
an
. 17 *
5
XX
IV.
26.
xxv.
20
mV
I. 4
1.:
XX
VII
. 27
Arte
rienb
lut.
Cur
are.
Art
erie
nbl. ;
darn
ach
succ
essi
v su
bcut
. In
ject
. vo
n
hrtc
ricn
llnt
wiil
irend
dcr
3Io
rplii
umna
rlios
c; 3
St.
Blu
t aus
d. V
. cav
a; g
leic
hz. m
it N
r. 67
geu
omm
en.
40 cg
3l
orph
in.
nach
Nr.
67.
Art
erie
nblu
t. B
lut g
leic
hz.
aus
dem
per
iph.
End
e dc
r V. j
ugul
ar.
Hie
rauf
int
rave
n. Iu
j. ei
ner
ster
il. P
yocy
an.-C
ultu
r. C
oma;
kei
ne T
empe
ratu
rerh
ohun
g; A
rter
ienb
lut.
Blu
t am
d.
peri
ph-E
nde
(1. V
. jug
ular
.; gl
eich
z.
mit
Nr.
72.
Art
erie
nblu
t; T
emp.
d.T
hier
cs 3
0° d
ar.
Inj.
v. 1
60B
Blu
t gl
eich
zeiti
g au
s de
r V. c
ava
1 COC
. IC
riim
pfe.
A
rter
ienb
lut
wah
rend
der
Coc
ainv
ergi
ftnng
; s/
4 St
.
Blu
t gl
eich
zeiti
g au
s de
r V
. ca
va;
Ten
iper
atur
d.
Art
erie
nblu
t; l
etzt
e Po
rt. e
iner
1et
alen
Hiim
orrh
agie
nach
74-
75.
Thi
eres
39.
9 O.
unm
ittel
bar
nach
76-
77
eing
clei
tet.
Art
erie
nbl;
Tem
p. d.
Thi
eres
39 - 6
$~;;C'?I;~;~;' B
lut
glei
chze
itig
aus
der
V.
cava
1 Pyocyan.-Cultur.
Art
erie
nblu
t; w
tihre
nd d
er P
yocy
aneu
sver
giftu
ng
Blu
t gl
eich
zeiti
g ai
ls d
er V
. ca
va.
Tem
pera
tur
d.
28/,
Stun
den
nach
79-
80.
Thi
eres
40.
6 O.

- 1 Numme:
p N N N Sauerstoi A 6 6 Volumen
w 01 w NNNll c procent
N Hamogl.
Q
?
L.
cc cc
WWW WW WWW OWW -am OlPW
??? ?? 000 PPN CnN CnulN 000 00 000
N
WWW a40 Olul
Y-'?
re- CnPP N-lW
ere wee WWP
Nummer
Procent Trocken- riickstanc
Procent Hiimogl.
Sauerstof totale Menge
Sauerstofi in Wasse:
geliist Sauerstofl v.Hlmogl gebunden Sauerstotl per 1006 Hiimogl.
-
-
L
0)
E z 96
97 5 -
-
98
99
100
101
102
103
0.18
41
0.15
94
-
-
c *g
g2
$3
&5j ri
13.5
4 14
.11
-
__
_
9.62
9.
58
15 * 7
5 15
32
16.2
2
13 * 5
2
0.50
23 '0
.139
9 0.
4704
10.
1293
-
~
u;;.: -3.2
%5
mc3
-
-
1.03
4 1
a035
1.02
3 1 *0
23
1.03
9 1
*038
1.
040
1 -0
34
0.42
07
0-41
87
0-49
99
0.39
78
-
-
* g$
-2
.S gJ
am -
-
0.35
9 0.
364
0.36
7 0
* 35
7
0.35
1 0.
362
0.38
0
0.36
4
0.11
99
0.11
55
0.13
16
0.11
00
-
-
5 $2
582
8% g
n*
17.8
2 17
.41
-
-
12.6
6 13
* 42
20 * 5
4 19
-69
21-4
1
17.3
4
-
-
SA
n.
."?
s gs
g
E%
g
& '2
nz -
-
131.
6 12
3.4
131.
6 14
0.1
130.
4 12
8.6
132.
0
128.
2
-
-
5 rt
: .9 .z*
$W
mg
ad
-
~
366.
5 33
9.0
358 *
7
392.
5
371.
6 35
5.6
347.
5
354 *
6
Hltm
oglo
bine
.
0.14
13
0.18
99
0.15
83
0.14
86
0.17
37
0.13
94
e z H
amog
lobi
n da
rges
tellt
aus
dem
Blu
te N
r. 38
7r
17
z
11
77
27
71
39
Ham
oglo
bin
darg
este
llt a
us d
em B
lute
Xr.
67
7,
77
71
:t
68
Hat
nogl
obin
dar
gest
ellt
aus
dem
Blu
te K
r. 2
9 77
77
17
77
31
;7
77
71
7.
32
Ham
oglo
bin
darg
este
llt a
us d
em B
lute
Kr.
53
M m 0 m Q P 9 E 8 ;
k E
71
77
c3
4
Q
77
71
17
72
+3 b

118 CHRISTIAN ROHR:
Die in vorsteheader Tabelle ent,haltenen Versuche iiber den specifischen Sauerstoffgehalt des Blutes wollen wir , der naheren Be- trachtung wegen, in folgende Gruppen eintheilen:
I. Versuche iiber normales Arterienblut aus verschiedenen Indi- viducn.
11. Verglciche zwisclien normalem, gleichzeitig demselben Indivi- duum cntnommenem Arterien- und Venenblut, wiihrend die Blutstriimung in den Gefassen nicht durch das Entnehmen der Proben gestiirt war.
111. Versuche iiber den specifischen Sauerstoffgehalk vor und nach eincm Blutverluste.
IV. Versuche uber die Verinderung der genannten Griisse durch Ein- athmung sauerstoffarmer Luft und durch Hemmung des Athem- zuges.
V. Versuche iiber den Einfluss verschiedener Giftstoffe auf den specifischen Sauerstoffgehalt (Curare, Morphin, Cocain, Pyocy- aneus-Cultur).
VI. Dicse Gruppe umfasst eine Zusammenstellung verschiedenartiger, vorzugsweise zur Orientirung fur fernere Un tersuchungen an- gestellter Versuche. Hierunter sind Versuche mit Venenblut, welches theils einem grijsseren abgesperrten Venengebiet, theils dem peripheren Ende durchschnittener kleinerer Venen entnommen ist, mit einbezogen. Ferner findet man in dieser Abtheilung Versuche uber die Veranderungen des Arterienblutes bei Sper- rung der Aorta thoracica, sowie einen ehzelnen Versuch mit Blut aus der Vena porta.
VII. I n dieser Abtheilung findet man einen Vergleich zwischen ver- schiedenen Blutproben und den aus diesen dargestellten Hamo- globinen.
Im Versuche XIX sind die Blutproben 57, 58, 59 unter sehr un- klaren Bedingungen entnommen. Die Proben 57 und 58 sind mittels eines hoch in die VeniL cava durch die Vena femoralis eingefiihrten Ka- theters entnommenes Venenblut. Die Aorta theracica war gesperrt und es wurde gleichzeitig eine Sperrung der Vena cava beabsichtigt oberhalb des Ortes, an dem die Probe genommen wurde, was aber missgliickte, und es ist also das Venenblut nicht einem gesperrten Venengebiet ent- nommen, sondern stammt vonugweise aus dem vorderen The& des K6r- pers, dessen Arteriensystem nicht abgesperrt war. Die Probe 59 sollte ihrer Absicht nach Arterienblut bei Aortssperrung sein, wedge Minuten vor Entnehmung der Probe schlug jedoch die Spermng fehl.
UERER DEN SPECIFISCHEN SAUERSTOFFGEHALT DES BLUTES. 1 19
-~
373 373 358 353 342 342 337 331 329 328 328
In den folgenden Special tabel len sind die Versuche mit Kum- mern versehen, die den Nummern der Generaltabelle entsprechen, deren Bezeichnungen fur die einzelnen Rubriken ebenfalls in den Special- tabellen angewendet sind, nur sind die Zahlen der Rubriken a,,= und ere, um Bruchzahlen zu vermeiden, mit 1000 multiplicirt. Ferner sind, wie in der Haupttabelle, die Resultate der Versuche niit Glan’s Apparat mit Cursivschrift gedruckt. In den Tabellen bedeutet A das Arterien-, V das Venenlilut.
180 481 137 568 167 466 279 791 271 792 315 923 273 813 288 871 271 826 Y59 789 273 833
I. N or m a 1 e s A r t e r i en 11 I u t. Die untenstehende Tabelle enthllt samm t l iche Versuche mit
normalem Arterienblut verschiedener Individuen.
Tabelle I. - ~
$&
8 8 G 5 go
*e
~ - 79 33 49 40 25 23 29 46 36 67 74
- ~
g G Y m i ;
6 ” a % ~ - 468 437 435 406 401 390 386 384 383 380 379
nor ‘* f e
- - Y
J2 qa ;I FO ;- j .9
49 56 65 58 51 60 74 74 68 63 62
.o
-- -
- __ $&
a $ 8%
$ - 60 70 38 27
8 48 21 19 64 1
62
- __ Y
v2 *a ; B
: 0 L?g 2 a - ” ._ - -
65 75 71 59 65 56 63 69 82 67 63
- - j m
:il = a a * 2.8
S
16 20 30 11 16 26 4 5 6
15 35
- -
Der speciEsche Eisengehait zeigt Verschiedenheiten von 328 bis 468; die grosste Abweichung ist also 180 auf 468 oder ca. 30 Proc. Die grosste Abweichung in a, (Eisenmenge pro Einheit absorbirten Liohtes) findet sich in den Versuchen mit dem Vierordt-Kriiss’schen Apparat, bei welchem das Maximum 180, das Minimum 127 ist. Die Abweichung also 53 auf 180 oder ca. 30 Proc.
Einen Ueberblick uber das Verhiltniss zwischen dem specfichen Sauerstoffgehalt und der Lichtabsorption des Blutes gewinnt man am besten durch eine graphische Darstellung, wie die umstehende. Es sind hier die laufenden Nummern der 22 Versuche der Tabelle lilngs der Abscissenaxe angefiihrt, als Ordinate sind aufgefihrt t h e 5 die
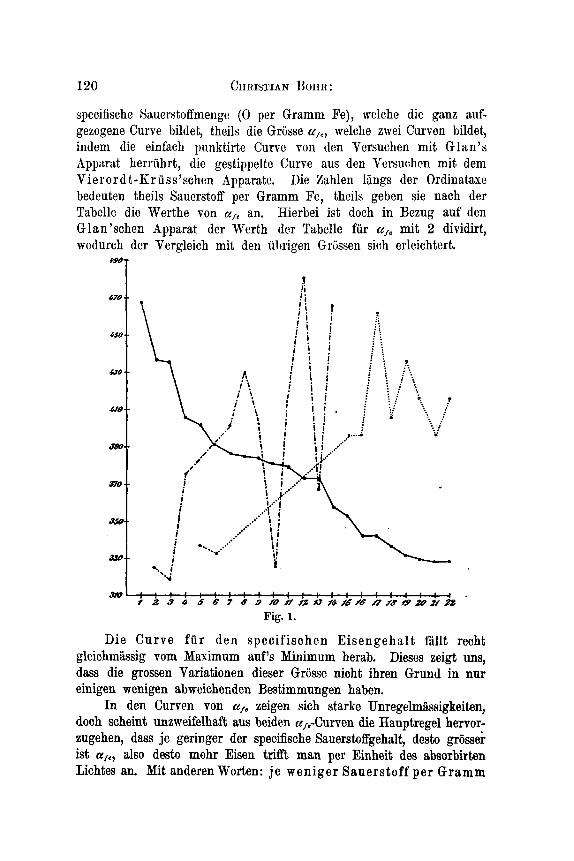
120 CHRISTIAN Horm:
specifische Sauerstoffmenge (0 per Gramm Fe), welche die ganz auf- gezogene Curve bildet, theils die Grosse a,., welche zivei Curven bildet,, indem die einfach punktirte Curve Ton den Versuchen mit Glan’s Apparst herriihrt, die gestippelte Curve aus den Vcrsuchcn mit dem Vierord t-Kriiss’schen Apparate. Die Zahlen liings der Ordinataxe bedeuten theils Sauerstoff per Gramm Fe, theils geben sie nach der Tabelle die Werthe von tc,, an. Hierbei ist doch in Bezug auf den Glan’schen Apparat der Werth der Tabelle fur a,, mit 2 dividirt, wodurch der Vergleich mit den iibrigen Griissen sich erleichtert.
490-
470. -
450.-
430.-
uo--
m- -
S70--
dzB-.
fl 2 3 4 5 6 7 8 9 l O f l ~ Z # 7 4 l 5 ~ 6 ~ 1 8 ~ 9 ~ 2 ~ 2 z w L : ; : : : : : : ! ! ! : : : : ! ! : : : : J .
Fig. 1.
Die Curve f u r den specifischen Eisengehal t fallt recht gleichmassig vom Maximum auf’s Minimum herab. Dieses zeigt m, dass die grossen Variationen dieser Grosse nioht ihren Grund in nur einigen wenigen abweicheEden Bestimmungen haben.
In den Curven von a,, zeigen sich starke Unregelmlssigkeiten, doch scheint unzweifelhaft aus beiden cc,,-Curven die Hauptregel hervor- zugehen, dass je geringer der specifische Sauerstoffgehalt, desto griisser ist ufe, also desto mehr Eisen trifft man per Einheit des absorbirten Lichtes an. Mit anderen Worten: j e weniger Sauerstoff per Gramm

i:
UEBER DEN SPECIFISCHIEN S AUERSTOFFGEHALT DES BLUTER. 12 1
Eisen, um so weniger Lichtabsorption per Gramm Eisen. BUS einer friiheren Abhandlung' crsehen wir, dass eben dieselbe Re gel f ii r H Cim o g 1 o b i n e v e r s c h i e d en e n S au e r s t o ffg e h a1 t s g i 1 t. Die Anschauung, zu der wir in der Einleitung hingefuhrt wurden, dass der verschiedene specifische Snuerstoffgehalt (0 per Gramm Eisen) seinen Grund in der verschicdenen Menge der ini Blute befindlichen Hiimoglobine niedrigeren oder hoheren Eisengehaltes habe, findet in dieser Weise durch die Untersuchung der Lichtabsorption im Blute ihre Bestiitigung. Wo im Fulgenden uns eine hinlangliche Anzahl von Versuchen zu Gebote steht, werden wir stets die obenstehende Regel riicksichtlich des Verhaltsnisses zwischen dem Sauerstoff, dem Eisen und der Lichtabsorption im Blute wieder finden; wo nur wenige Ver- suche uns zll Gebote stehen, bekommen wir uber diese Frage nichts zu wissen wegen der uberall auftretenden Ausnahmen von der Regel. Die moglichen Ursachen dieser Abweichungen sind in der Einleitung zu dieser Abhandlung hinlanglich abgehantlelt worden (S. 107).
Res um6. I n dem normalen Arter ienblute aus verschiedenen I n d i -
viduen gleicher Art ist der specifische Sauerstoffgehslt des Blntes sehr variabel.
11. Normales Arter ien- und Venenblut. Die Tabelle auf S. 122 umfasst Versuche von finf verschie-
denen Individuen. Die Arterienblutproben sind der A. carotis oder femoralis entnommen, das Venenblut gleichzeitig aus dem untersten Theile der V. cava gerade iiber der Theilung derselben in V. ilacae und auf folgende Weise: ein gewbhnliches elastisches Katheter wurde in die eine V. femoralis binein und in die V. cava bis kurz iiber den Theilungsort hinaufgefuhrt ; mittels des Katheters wurde darauf lang- Sam mit Hulfe einer Spritze eine Probe entnommen. In diesen Ver- suchen wird also Arterienblut mit frei stromendem, grasstentheils aus den Muskelvenen herruhrendem Venenblute verglichen.
Der specifische Eisengehalt ist uberall im Venenblute niedriger, als im cntsprechenden Arterienblut , die Unterschiede sind bezw. 48, 28, 20, 13 und 7, und (siehe die Tabelle) desto grosser, je grosser der absolute Werth des specifischen Eisengehaltes ist. Perner ist zu bemerken, dass die procentische Eisenmenge in keinem der Versuche im Arterien- und im Venenblute dieselbe ist, und ebenfalls in keinem
Ueber die Verbindungen des Hamoglobins mit Sauerstoff. S . 8'1 f.
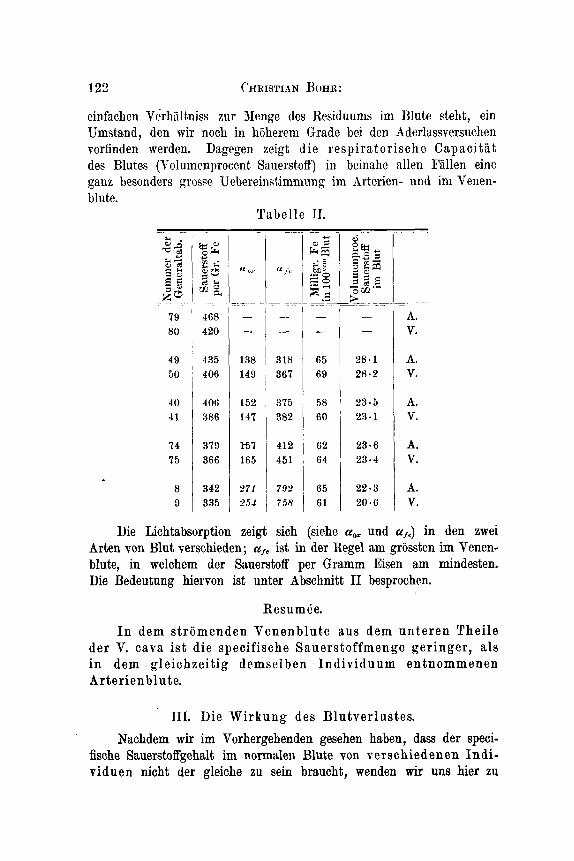
122 cHRISTIAN HOHR:
40 I 406 1 152 1 375 41 386 147 382
74 379 t57 412 75 366 165 451
8 342 271 7931 9 335 2551 1 758
cinfachcn Vcrhlilt8niss zur Nenge des Residuums im Blute steht, ein Umstand, den mir noch in hoherem Grade bei den Adorlassversuchen vorfinden werden. Dagegen zeigt die respiratorische Capacitat des Blutes (Volumenproccnt SauerstofT) in beinahc allen Fiillen eine ganz besonders grosw Uebereinst,immung im hrterien- und in1 Venen- blute.
Tabelle 11.
58 ~ 2 3 . 5 A.
, 64 ’ 2 3 . 4 v. 65 2 2 . 3 A. 61 20 .6 v.
80 1 420 V.
28.1 A.
Die Lichtabsorption zeigt sich (siehe u, und u,J in. den zwei Arten von Blut Tersehieden; ist in der Regel am grossten im Venen- blute, in welchem der Siuerstoff per Gramm Eisen am mindesten. Die Bedeutung hiervon ist unter Abschnitt I1 besprochen.
Resum Be. I n dem stromenden Venenblute aus dem unteren Thei le
der V. cava i s t die specifische Sauerstoffmenge ger inger , als i n dem gleichzeitig demselben Indiv iduum entnommenen Art e r i en bl u t e.
111. Die Wirkung des Blutverlustes. Nachdem wir im Vorhergehenden gesehen haben, dass der speci-
fische Sauerstoffgehalt im normalen Blute von verschiedenen Indi - viduen nicht der gleiche zu sein braucht, wenden wir uns hier zu

UEBER DEN SPECIFISCHRN SAUERSTOFFGEHALT DES ELUTES. 1 23
Versuchen, in denen einer Verlnderung der genannten Grosse bei ein em u n d dems elb en Individuum nachgestrebt ist.
Das Aderlassen hat sich als gutes Mittel erwiesen, um derartige Veranderungen hervorzurufen. Die Wirkung desselben ist an im Ganzen neun Individuen versucht worden. Bei acht derselhen, yon denen einige mehrere Male gepriift wurden, trat stets eine Wirkung uberein- stimmender Art zu Tage, ein einziges Thier nur liess sich nicht von Aderlksen beeinflussen; es wurden im Ganzen acht Bestimmungen mit Blutproben dieses Individuums angestellt ; in ihneu dlen zeigte sich das Hamoglobin so gut wie unveriindert.
Die Versuche betreffen im Wesentlichen nur das Arterienblut; der Weise nach, in der sie angestellt wurden, zerfallen sie in zwei Gruppen. In der ersten hat man, nachdem eine Blutprobe zur Untersuchung entnommen und, wenn nijthig (bei grosseren Thieren), gleichzeitig ein fernerer Blutverlust hervorgerufen worden ist, das Thier in einigen Tagen sich selbst uberlassen, worauf man von neuem eine Untersuchung vorgenommen hat; in einem einzelnen Falle hat man die Bestandtheile des Blutes in einem Monate sich regeneriren lassen und dann auf's Neue das Blut untersucht. Diese Versuche sind in der Tabella I I Ia zusammengestellt worden. In einer zweiten Gruppe von Versuchen ist, nachdem die erste Blutprobe entnommen war und das Thier noch einen Theil Blut mehr verloren hatte, ein dem Blutverluste entsprechen- des Quantum von 0.7 procentiger ClNa-Losung von 38O injicirt worden. Nach dem Verlaufe einer halben oder ganzen Stunde ist darauf eine neue Blutprobe zur Untersuchung herausgenommen, womit der Versuch abgeschlossen wurde. (Diese Versuche sind in der Tabelle I I Ib ent- halten.)
Mit Ausnahme der vom Hunde Nr. I1 stammenden Versuche Nr. 8-15, in denen sich der specifische Sauerstoffgehalt im Arterien- blute uberall ungeahr unveriindert gehalten hat oder wenig gestiegen ist, hat sonst der Blutverlust in sammtlichen Versuchen ein bisweilen sehr bedeutendes Fallen der genannten Grosse verursacht.
Gleichzeitig (mit Ausnahme der Versuche 29-30) ist der Werth von u,, gestiegen, bisweilen sogar in so bedeutendem Grade, dass a, sich unveriindert gehalten hat, wie es auch i n gewissen F l l l e n beim Uebergang einer Hamoglobinlosung von der y- zur @-Nodification ge- schieht.' In der Regel Gllt doch auch a, (Sauerstoff per Einheit ab- sorbirten Lichtes) durch Blutverlust, wie unter anderen es die Versuche 16-19 zeigen, in denen man ferner beobachtete, dass die Veriinde-
~
Ueber die Verbindungen des Hamoglobins mit Sawstoff. S. 88.

124 CHRISTIAN HOHR:
rungen bei der Regeneration des Blutes in einer IEichtung gehen, die denen entgegengesetzt ist , welche durch Blutverlust hervorgerufen werden.
Die Unterschiede in dem specifischen Sauerstoffgehalte, die zwischen den Blutproben 29-31 beobachtet worden sind, sind im Ungefihren zwischen den von ihnen darges te l l ten Himoglobinen wieder ge- fundcn worden (siehe den Abschnitt VII).
Wie schon im vorigen Abschnitte beriihrt, ist die Eisenmenge im Blute vor und nach dem Aderlass nicht stets der Menge des Resi- tluums (siehe z. B. Nr. 1 und 6) proportional.
- $ b cd
9% $?I
- R a.
Y
16 17 18 19
1 4 6
8 9 10 11 12 14
- - -~
;c Y .z a '+a g a a 2;; 4 2 . e- 23.2 19.0 16.0 21.2
20.7 16.1 13.3
20.9 20.4 16.1 16.4 15.9 17.1
- __
%g g& *
2 - m %
~ ._
- 316 318 331
328 316 304
342 335 343 348 354 346
Tabel le IIIa. - __
ffo,
- 288 249 258 " 8
259 270 274
?71 ?54 506 26'3 "77 ?XO
-- -~
.. .
- 787 812 871
789 856 924
792 758 893 756 781 807
A. normal A. 4 Tage nach Nr. 1 A. 8 11 11 11 1
"'1 normal V. "') 6 Tage nach Nr. 8 V. A. 12 Tage nach Nr. 8 A* 16 11 11 11 8
Die Griisse des Blutver lustes , welche nothwendig ist, um die ofter genannten Veriinderungan im Blute hervorzurufen, ist nicht gross. In keinem unserer FHlle hat der Blutverlust die HHlfte .der Blutmenge uberstiegen, ein Blutverlust von einem Fiinftel der Blutmenge hat in Nr. 21-22 eine starke Wirkung hervorgerufen, und eine solche ist in einem anderen Versuche noch deutlich bei einem Verluste von einem Siebentel der Blutmenge (siehe Nr. 29,31). Indess muss man hier darauf vorbereitet win, grosse individuelle Unterschiede anzutreffen , wie sie uns die Untersuchung des Hundes Nr. II liefert. Die Reihe von Ver- suchen, die mit diesem Thiere angestellt worden sind, bietet uns frei-
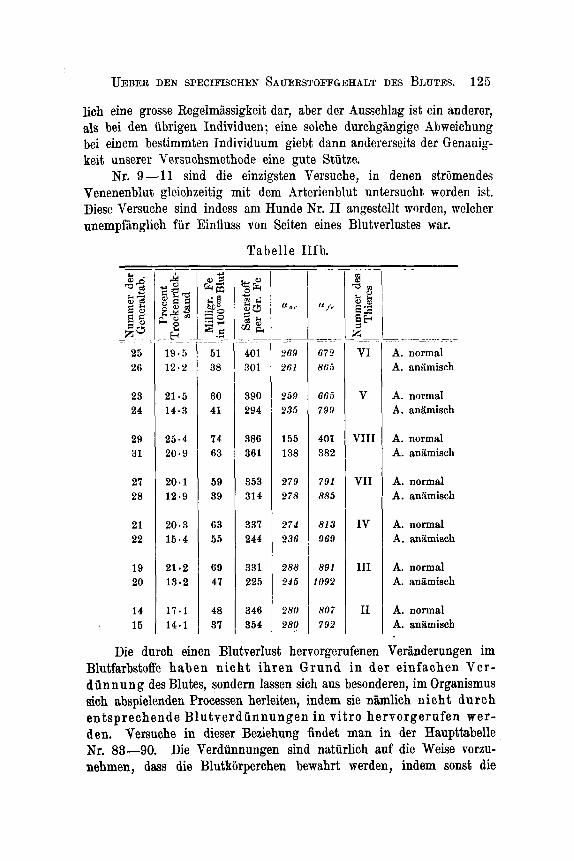
UEBER DEN SPECIFISCHEN SAUERSTOFFGEHALT DES HLUTES. 125
lich eine grosse Regelmhsigkeit dar, aber der Ausschlag ist ein anderer, als bei den ubrigen Individuen ; eine solche durchgangige Abweichung bei einem bestimmten Individuum giebt dann andererseits der Genauig- keit unserer Versuchsmethode eine gute Stutze.
Nr. 9-11 sind die einzigsten Versuche, in denen striimendes Venenenblut gleichzeitig mit dem Brterienblut un tersucht worden ist. Diese Versuche sind indess am Hunde Kr. I1 angestellt worden, welcher unempfiinglich fur Einfluss von Seiten eines Blutverlustes war.
Tabelle IIIb. - ~
$ & = $ 53 g s s o E W
25 26
23 24
29 31
27 28
21 22
19 20
14 15
- -- -* S 2 Z
E J k - .~
E
@ Z
19.5 12.2
21.5 14.3
25.4 20.9
20.1 12.9
20.3 15.4
21.2 13.2
17 .1 14.1
- __ 0
G.”iijl &$ 22 .“ 0 =0
.+ ~- ._ ~
51 38
60 41
14 63
59 39
63 55
69 47
48 37
- ~
2G p 2 k
Y m J
. _
401 301
390 294
386 361
353 314
337 244
331 225
346 354
- _-
a o r
.__ _. ~
969 261
259 235
155 138
279 278
274 236
288 245
280 280
- __
n ye
.
672 865
666 799
401 382
791 885
81 3 969
891 1092
807 792
~
A. normal A. anlmisch
A. normal A. anamisch
A. normal A. aniimisch
A. normal A. aniimisch
A. normal A. aniimisch
A. normal A. anamisch
A. normal A. aniimisch
Die durch einen Blutverlust hervorgerufenen Veriinderungen im Blutfarbstoffe haben n i ch t ih ren Grund in der einfachen Ver- di innung des Blutes, sondern lassen sich aus besonderen, im Organismus sich abspielenden Processen herleiten, indem sie niimlich n ich t durc h entsprechende Blutverdunnungen in vi t ro hervorgerufen wer- den. Versuche in dieser Beziehung findet man in der Haupttabelle Nr. 83-90. Die Verdunnungen sind naturlich auf die Weise vorzu- nehmen, dass die Blutkorperchen bewahrt merden, indem sonst die

126 CHRISTIAN BOHR:
383 321
358 331
406 386 388 389
Concentration des Hlmoglobins verandert wird. Bei einer dreimaligen Verdunnung mit 0.7 procentigem ClNa zeigte sich in einem Falle (Nr. 88-89) nur ein Abfall von 149 auf 147ccm Sauerstoff per 1006 Hlmoglobin; bei eincr achtmaligen Verdunnung mit ClNa (Nr. 83-84) eine Steigerung der Sauerstoffaufnahme von 114 auf 118 ccm; wurde das- selbe Blut dagegen mit procentiger NazCO,-Losung verdunnt, wodurch d a Hlmoglobin in Aufliisung gerieth, so wurde in bedeutendem Grade mehr Sauerstoff (133 ccm) aufgenommen, wahrscheinlich weil das Hamo- globin nun in weit geringerer Concentration vorhanden war, als zur Zeit, da es im Stroma der Blutkorperchen eingeschlossen war.'
Re sum i: e. Durch Blutver lust n immt im Arter ienblute der speci-
fische Sauerstoffgehal t des Blutes ab.
154 131
167 167
152 147 144 160
IV. Einathmung sauerstoffarmer Luft und Hemmung der A t hm un g.
Bei den in der Tabelle IVa angefuhrten Versuchen entnahm man zuerst dem Thiere eine Blutprobe und liess dieses darauf 1/4-1/z Stunde durch ein Klappenventil aus einem grossen Spirometer ca. 9 Procent
Tabelle IVa. - $d
$3 6 2 I$
d
- __ 36 37
38 39
40 41 42 43
- - __
n/e
- __ 402 407
466 505
375 382 371 411
~
A. normal A. Einathmung v. 8.6 Proc. 0
A. normal A. Einathmung v. 8.9 Proc. 0
"*) V. normal
Einathmung $:) von 8.3 Proc. saueratoff
Sauers t off en t h a1 t ende Luft einathmen; wihrend des Einathmens der sauerstoffarmen Luft aurde darauf auf's Neue eine Probe Blut zur Untersuchung genommen.
Ueber den Einfluss Liner Concentratimsaeriinderung auf die Sauerstoff- aufnahrne siehe: Ueber die Verbindmg des Hamglobins mit Saws to f .

URBER DEN SPECIFISCHEN SAUERSTOFPGEHALT DES BLUTES. 1 27
Im Arterienblut ist der specifische Sauerstoffgehalt durch Ein- athmen sauerstoffarmer Luft geringer geworden. Die Eisenmenge hat sich indess so gut wie constant gehalten; die respiratorische Capa- citat (Volumenprocent Sauerstoff) ist also verandert.
In den Nummern 40-43 ist gleichzeitig Arterienblut und das stromende Venenblut aus der Venn cava. untersucht (wie im Ab- schnitt 11). Wiihrend der specifische Sauerstuffgehalt im Arterienblut von 406 auf 388 herabgegangen ist, ist derselbe im Venenblut so gut wie unveriindert geblieben (386-359).
Hinsichtlich der Lichtnbsorption, die weder hier noch bei den anderen Versuchen constant ist , liisst sich aus unseren Bestimmungen keine besondere Regel herleiten.
Aus den Blutproben Nr. 38 und 39 sind Hlmoglobine dargestellt worden; diese zeigen (siehe Abschnitt VII) den gleichen Unterschied des specifischen Sauerstoffgehaltes als die Blutproben. (Die DilTerenz zwischen den Blutproben Nr. 38 und 39 ist 27, zwischen den ent- sprechenden Hamoglobinen 28.)
Die Tabelle IVb enthalt zwei Versuche iiber den Einfluss des Ers tickens auf die hier besprochenen Blutrednderungen. Im ersten Falle wurde die erste Blutprobe wahrend der Erstickungskrlmpfe genommen, die zweite funf Minuten spater, nachdem die Athmungs-
Tabelle IVb.
47
hindernisse entfernt waren, wahrend das Thier tief und ivhig athmete. In dem zweiten Versuche wurde zuerst eine Probe normalen Arterien- blutes genommen, darauf wurde die Trachea des Thieres versperrt und unmittelbar vor Eintritt des Todes wurde eine Probe des Erstickungs- blutes genommen.

128 CHRISTIAN BOHR:
Re sum4 e. 1) Durch Einathmen saucrstoffarmer Luft fa l l t der spe-
c i f i s c h e S au e r s t o f f g e h a1 t d e s A r t e r i e n b lu tes w i h r e n d d e r des Venenblutes sich unverandert halt.
2) I m Erst ickungsblute i s t der specifische Sauerstoff- geha l t vermehrt.
V. Ve r g i f t un g e n. I n Bezug auf die in nebenstehender Tabelle V enthaltenen Ver-
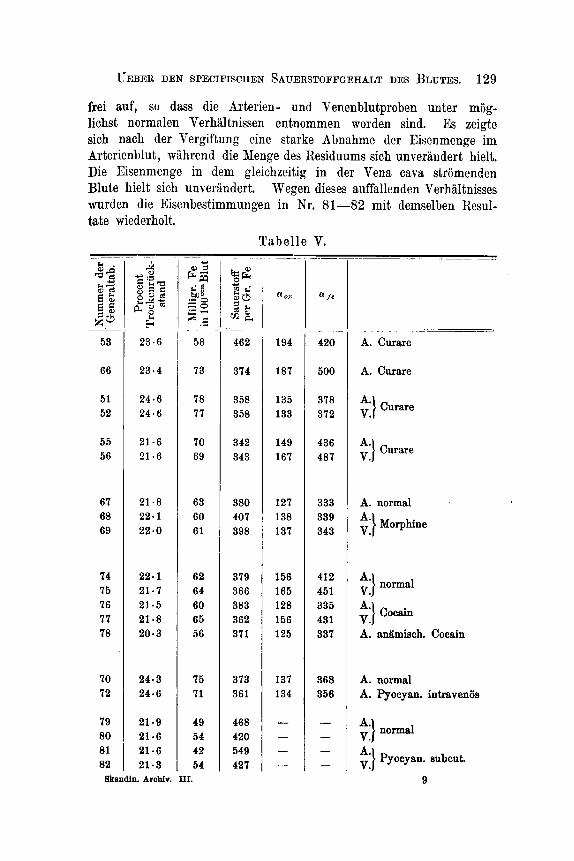
suche ist Folgendes zu bemerken: I m Curareversuche Nr. 53 war die kiinstliche Respiration wegen einer in den Ventilen entstandenen Undichtigkeit sehr unvollkommen. Im Morphinversuche bekam das Thier, welches 2Gkg wog, subcutan im Verlaufe von drei Stunden im Ganzen 40 cg Morphin, darauf wurden Proben des Arterien- sowie des stromenden Venenblutes der Vena cava sntnommen. Das zum Cocainversuche angewandte Thier wog 42 kg und bekdm subcutan 16cg Cocnin: es traten starke Krampfe auf, die Temperatur stieg von 39.6 auf 39.9O. Nachdem die Krampfe aufgehort hatten, wurden die Proben Nr. 7 6, 77 genommen (Arterienblut und gleichzeitig stromendes Venenblut aus der Vena cava). Darauf wurde das Thier durch Ver- blutung getiidtet. Das zuletzt ausstromende Blut wurde aufgefangen und analysirt (Nr. 78); es enthielt nur 56 Proc. Eisen. Das zuerst entleerte Blut dagegen GO Proc. Damit iibereinstimmend zeigte diese letzte Blutprobe (Nr. 78) eine Veranderung des specifischen Sauerstoff- gehaltes wie bei Anamie (siehe sub 111). Bei der eigentlichen Ver- g h n g ist der specifische Sauerstoffgehalt so gut wie unverandert ge- blieben, sowohl im Venen- als im Arterienblute, dagegen zeigt letzteres, n ioh t aber das gleichzeitig i n der Vena cava s t r i imendevenen- blut , eine starke Veranderung der Lichtabsorption, indem cc, von 156 auf 128 gefallen ist. Die Richtigkeit dieses auffallenden Resultates, die vorliiufig als nicht erklarbar hinstehen muss, wird dadurch an- gezeigt, dass es sich genau in der zweiten Probe von Cocain-Arterien- blut (Nr. 78) wiederholt, obgleich die beiden Proben ganz unabhlngig von einander untersucht wurden. I n Bezug auf die Pyocyaneus- Vergiftungen wurde in Nr. 70, 72 eine griissere Menge sterilisirter Pyocyaneus-Cultur in das Venensystem des Versuchsthieres eingespritzt; es folgte ein comatoser Zustand, jedoch keine Temperaturerhohung. In den Versuchen Nr. 79-82 wurde einem anderen Thiere successive 10 wm einer neuen sterilisirten Pyocyaneus-Cultur subcutan injicirt. Die Tem- peratur stieg von 39.6 auf 40.69 Das Thier, wahrend des ganzen Versuches ungebunden, stand wiihrend des Entnehmens der Proben
11 *
lp
2
% g
g
ll I,
mw
w
ww
l
pw
w
wlp
wlp
u-
Q
2
4-
cn
l-
N
ww
w
4u
J
NO
D
0 0
WC
OW
cn
l-
w
w
II
II
g
g
? C? &?
C??
-
C?
s c -
d
(D
C?
5
-4c P (D

130 CIIRISTIAN BOHR:
Die Verinderungen, welche das Blut durch die Uorphinvergiftung erlitten hat, findet man in den aus Nr. 67 und 68 dargestellten Himo- globinproben wieder (siehe Abschnitt 111). Wahrend der Unterschied im specifischen Eisengehalt in Bezug auf die Blutproben 27 .war, war er in Bezug auf die Himoglobine 34.
Einige Tersuche (siehe die Haupttabelle Kr. 91-95) sind iiber den Einfiuss angestellt, welchcn das Norphin! das Cocain und eine sterilisirte Pyocpneus-Cultur auf das Blut in vitro bei 38 O ausiibt,. Die beiden erstgenannten Gifte zeigten sich ohne irgend einen Einfluss auf die Sauerstoffaufnahme dcs Ulutes; diejenige Verinderung im speci- fischen Eisengchalte, die oben Bei der Vergiftung des Thieres nachgewiesen worden ist, hat also ihren Grund nicht in irgend einer directen Einmirkung auf das Blut. Dagegen hat das Pyocyaneusgift in vitro dem Blute einen specifisch hiiheren Sauerstoffgehalt gegeben. Es bedarf dieses jedoch einer naheren Untersuchung, nicht so sehr, weil der Versuch nur ein vereinzelter ist, als weil ich es versiiumt habe, zu untersuchen, ob die zugesetzte Pyocyaneus - Cultur den Sauerstoff dissociabel band. In einer folgenden Arbeit wird deshalb diese Frage zur erneuerten Behandlung aufgenommen werden.
Re s um 6 e. Bei der Curarcvergiftnng i s t der normale Unterschied an
s p e ci fi s c h e m S a u e r s t o f f g e h a 1 t (s i e h e A b s c h n i t t II) z w i s c h en dem Ar ter ienblu t und dem gleichzeitig im unteren Theile der Vena cava stromenden Venenblute verschwunden; das Arter ien- und das Venenblut sind hier gleich.
Bei der Morphinvergiftung steigt im Arter ienblut der specifische Sauerstoffgehalt, das Arterien- und Venenblut zeigt in obengenannter Hinsicht Verschiedenheiten, die in derselben R ich tung gehen, wie es normal der Fa l l ist.
Bei der Cocainvergiftung wird der specifische Sauerstoff- geha l t nu r wenig vergndert , d a er indess e in wenig im Arter ienblute s te igt und im Venenblute s ink t , wird der Unterschied zwischen dem stromenden Arter ien- und Venen- blute grijsser nach der Vergiftung (211, als vor derselben (13). Die Lichtabsorption v e r l n d e r t sich im Arter ienblute und im Wesentlichen n u r in diesem.
Bei der Pyocyaninvergiftung wird das Venenblut n u r wenig verandert, der specifische iSauerstoffgehalt s te igt da- gegen s t a rk i m Arterienblute. Dadurch wird der Unterschied

UEBER DEN SPXCIFISCHEN SAUERSTOFFGEHALT DES BLUTES. 131
53 54
33
zwischen s t romendem Arter ien- und Venenblute weit grosser nach a l s For d e r Vergiftung.
23.6 58 462 194 420 A. Curare 22.2 66 376 213 565 V. Obturation der V. c8va
22.6 56 437 141 324 A. normal
VI. Gemischte Versuche. Die Tabelle VIa enthalt zmei Yersuche uber die Verinderung des
Blutes bei Sperrung eines griisseren Venengebietes. In beiden Fiillen zeigt sich im abgesperrten Venenblute ein starkes Fallen des speci- fischen Sauerstoffgehaltes; uoz steigt ein wenig oder bleibt unverandert. In Nr. 34 wurde nach Aufhiiren der Sperrung ein Aderlass vorge- nommen und darauf wiederum eine Probe des Arterienblutes ent- nommen. Dieses zeigt, wie alle hamorrhagische Versuche, ein Fallen des specifischen Sauerstoffgehaltes, wiihrend u,, steigt. Besonders be- merkenswerth ist bei diesen Versuchen das Verhiltniss zwischen dem Eisengehalt des Blutes und der Menge des Residuums; durch Sperrung des Venengebietes halt das Residuumsprocent sich ungefahr unverandert im abgesperrten Blute, wahrend der Eisengehalt desselben stark steigt. In Nr. 35 ist durch Aderlass die Menge des Residuums gefallen, jedoch die Eisenmenge im Arterienblute behalt die Grosse, welche sie beim Beginn des Versuches hatte. Sowohl in Nr. 33 als 35 sind doppelte Eisenbestimmungen ausgefiihrt worden, die genau iibereinstimmten.
34 23.8 35 I 17.3
Tabelle VIa.
70 344 146 408 V. Obturation der V. cava 55 327 146 448 A. anlimiach
In der Tabelle VIb sammeln sich alle die Versuche, die einen Vergleich erlauben zwischen Arterienblut und Blut aus dem peripheren Ende einer abgeschnittenen kleineren Vene. Hier ist uberall der spe- cifische Sauerstoffgehalt grosser im Venenblut als im Arterienblute. In Uebereinstimmung mit den Hlmoglobinversuchen (s. unter Ab- schnitt I) ist dagegen (die Eisenmenge per Einheit absorbirten Lichtes) geringer im Venenblute.
9'
132 CHRISTIAN BOHR:
386 389
373 383
328 373
316 337
304 326
155 401 148 380
137 368 137 356
259 789 ?94 787
27U 856 262 776
274 924 266 815
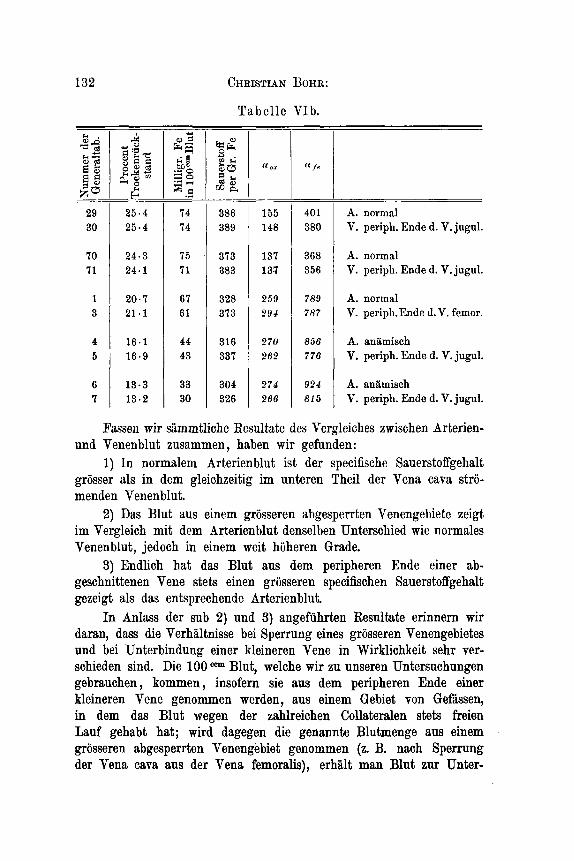
Tabelle VIb.
29 30
70 71
1 3
4 5
6 7
25.4 74 25.4 74
24.3 75 24.1 71
20.7 67 21.1 61
16.1 44 16.9 43
13-3 33 13.2 30
A. normal V. periph. Ende d. V. jugul.
A. normal V. periph. Ende d. V. jugul.
A. normal V. periph.Ende J.V. femor.
A. anlmisch V. periph. Ende d. V. jugul.
A. anamisch V. periph. Ende d. V. jugul.
Fassen wir sammtliche Resultate des Vergleiches zwischen Arterien- und Venenblut zusammen , haben wir gefunden:
1) In normalem Arterienblut ist der specifische Sauerstoffgehalt griisser als in dem gleichzeitig im unteren Theil der Vena cava stro- menden Venenblut.
2) Das Blut aus einem grosseren abgesperrten Venengebiete zeigt im Vergleich mit dem Arterienblut denselben Unterschied wie normales Venenblut, jedoch in einem weit hoheren Grade.
3) Endlich hat das Blut aus dem peripheren Ende einer ab- geschnittenen Vene stets einen griisseren specifischen Sauerstoffgehalt gezeigt als das entsprechende Arterienblut.
In Anlass der sub 2) und 3) angefuhrten Resultate erinnern wir daran, dass die Verhaltnisse bei Sperrung eines grosseren Venengebietes und bei Unterbindung einer kleineren Vene in Wirklichkeit sehr ver- schieden sind. Die 100 Blut, welche wir zu unseren Untersuchungen gebrauchen , kommen , insofern sie aus dem peripheren Ende einer kleineren Vene genommen werden, aus einem Gebiet yon Gefmsen, in dem das Blut wegen der zahlreichen Collateralen stets freien Lauf gehabt hat; wird dagegen die genannte Blutmenge aus einem grosseren abgesperrten Venengebiet genommen (2. B. nach Sperrung der Vena cava aus der Vena femoralis), erhalt man Blut zur Unter-
Proc
ent
g g
11 Sauer
stof
f 10
per
Gr.
Fe
I/ a
2 p 11
R
Oh
:
ch
(IN
umm
er d
er
Lu
O
1 Gen
eral
tab.
7 7
!Tm
cken
ruck
Pr
ocen
t
,,, I,
Illil
ligr.
Fe
* u1
in 1
oocc
m"1
31ui
-
,
stan
d 4
w
I
10
w
g
Saue
rsto
ff
* CD
w
w
pe
rGr.
Fe
410
??
??
~l
134 CHRISTIAN BOHR:
nicht der Fall mit den Unterschieden im Verhaltnisse zwischen der Lichtabsorption, dem Sauerstoff und dem Eisen. Die miiglichen Grunde dafur wurden in der Einleitung genannt.
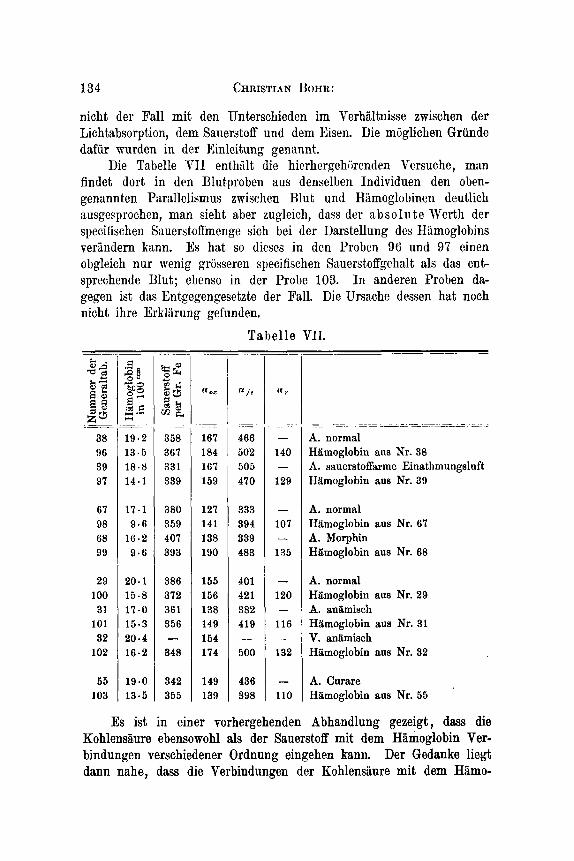
Die Tabelle TI1 enthiilt die hicrhergehijrenden Versuche, man findet dort in den Blutproben aus denselben Individuen den oben- genannten Parallelismus zwischen Blut und Hiimoglobinen deutlich ausgesprochen, man sieh t aber zugleich, dass dcr a b s o In t e Werth der specifischen Sauerstoffmenge sich bei der Darstellung des Hiimoglobins verindern kann. Es hat so dieses in den Proben 96 und 97 einen obgleich nur wenig griisseren specifischen Sauerstoffgehalt als das ent- sprechende H u t ; ebenso in der Probe 103. In anderen Proben da- gegen ist das Entgegengesetzte der Fall. Die Ursache dessen hat noch nicht ihre Erkliirung gefunden.
Tabel le VII. =
$& 5 E 3s 8 % g g
.~ z!? 38 96 39 97
67 98 68 99
29 100 31 101 32 102
55 103
- - c .- 0 $: 98 a ” - :Cd G
b . r *- w -. _ -
19.2 13.5 18.8 14.1
17.1 9.6 16.2 9.6
20.1 15.8 17.0 15.3 20.4 16.2
19.0 13.5
- __
%$ 20 i3g L 9 ; ;
~~ - -
358 367 33 1 339
380 359 407 393
386 372 361 356
348
342 355
-
- __
f f o r
-_ - 167 184 167 159
127 141 138 190
155 156 138 149 154 174
149 139
- __
/e
__ 466 502 505 470
333 394 339 483
401 42 1 382 419
500
436 398
-
I
~- . - 140
129 -
- 107
135 -
- 120
116
132
-
-
- 110
_ _ - - ~~ __ ____ -~ -.
A. normal Hiimoglobin aus Nr. 38 A. sauerstoffarme Einathmungsluft Hamoglobin aus Nr. 39
A. normal Hamoglobin aus Nr. 67 A. Morphin Hamoglobin aus Nr. 68
A. normal Hamoglobin aus Nr. 29 A. anlmisch Hiimoglobin aus Nr. 31 V. anamisch Hiimoglobin aus Nr. 32
A. Curare Hihoglobin aus Nr. 55
Es ist in einer vorhergehenden Abhandlung gezeigt, dass die Kohlensiiure ebensowohl als der Sauerstoff mit dem Hamoglobin Ver- bindungen verschiedener Ordnung eingehen kann. Der Gedanke liegt dann nahe, dass die Verbindungen der Kohlensaure mit dem Hiimo-

UEBER DEN SPECIFISCHEN SAUERSTOFFGEHALT DES BLUTES. 135
globin im circulirenden Blute ebensowohl gesetzmassig variiren als die Oxyhimoglobine. Indess lassen sich aus den in dcr Generaltabelle an- gefiihrten Data uber die Nenge der Kohlensaure im Blut aus dem Grundc keinc sichere Schlusse ziehen, weil man die Kohlensaure im Gegensatze zum Sauerstoffe an verschiedene zahlreicher rorhandene Snbstanzen im Blute gebunden findet.
Dagegen kunnte miiglicher Weise auf einem anderen Wege die Existenz verschiedener Kohlensiurehamoglobine gepriift werden ; ich hoffe spAter Untersuchungen hieriiber vorlegen zu kijnnen, sowie auch iiber das nihere Detail beziiglich des Verhaltnisses zwischen dem spe- cifischen Sauerstoffgehalt des Arterien- und des Venenblutes.
Zweites Capitol.
I. Ucber die Bedeutung des specifischen Sauerstoffgehal tes f u r die Sauerstoffspannungen i m Blute.
Im vorhergehenden Capitel haben wir in einer Reihe von Bei- spielen gesehen , dass der specifische Sauerstoffgehalt des Blutes con- stant ist. Um zu verstehen, welchen Einfluss dieser Umstand auf die Sauerstoffspannungen im Blute gewinnt, stellen wir uns zwei Blut- proben gleicher Temperatur A und B vor, welche beide gleichviel Ha- moglobin enthalten und jede fur sich ein gleich grosses Quantum Sauerstoff absorbirt haben , in denen aber der specifische Sauerstoff- gehalt verschieden ist. Gesetzt die letztgenannte Griisse wire am kleinsten in B, mit anderen Worten, gesetzt, dass B bei einem ge- gebenen Sauerstoffdrucke weniger Sauerstoff als A absorbire; es ist dann einleuchtend, dass die Sauerstoffspannungen in unseren beiden Blutproben versohieden sein miissen , obgleich pie beide gleichviel Sauerstoff per Gramm Hamoglobin enthalten und specie11 , dass die Spannung am grcissten in B sein muss, in der der specifische Sauer- stoffgehalt am kleinsten ist. In einer Blutprobe wird dann, wenn alles iibrige gleich ist, die Spannung wachsen, wenn der specifische Sauer- stoffgehalt abnimmt.
Wir haben ferner im vorigen Capitel gesehen, dass der specifische’ Sauerstoffgehalt in verschiedenen Gefhsgebieten eines und desselben Thieres verschieden sein kann und es auch hiufig ist, und dass der- selbe sich an einem lndividuum verandern lasst durch Eingriffe (Ader- l i s e , Giftstoffe), die in vitro keinen Einfluss auf das Blut ausiiben. Wir sind deshalb genothigt, die Veranderungen des specifischen Sauer- stoffgehaltes als Resultate besonderer Processe in den Geweben des Organismus zu betrachten.

136 CHEISTIAN BOHR:
Aus dem Obenstehenden leiten wir folgende fur die Lehre von der Spannung der Luftarten michtige Slitze ab.
Die Sauerstoffspannungen im Blu te sind bei gegebener Temperatur n ich t abh5ngig allein von der blenge der in1 B lu te per Gramm Hamoglobin lose gebundenen Sauerstoffes; durch eine Veriinderung des specifischen Sauerstoffgehaltes vermag n iml i ch das Gewebe des Orgnnismns einer gegebcnen Menge in dem Blute absorbir ten Sauerstoffs verschiedene Spannung zu geben, und specie11 eine desto griissere Span- nung, j e ger inger der specifische Sauerstoffgehalt ist.
Die Annahme liegt nahe, dass solche Spannungsveriinderungen zur Regulirung des Luftwechsels dient, dime Annahme werden wir im Folgenden bestitigt finden.
11. Ueber das Verhal tniss zwischen den Saucrstoffspannungen im Blu te und der Nenge von Sauerstoff, die im Plasma auf- gelijst, in jedem Augenblick zur Disposition der Gewebe-
zellen steht. Wie wir im vorigen Abschnitte andeuteten, sind die Variationen
in dem specifischen Sauerstoffgehalt ein Mittel zur Regulirung der Sauer- stoffspannungen in1 Blute; bevor wir im Polgenden dazu iibergehen, die Art dicser Regulirung durch Beispiele unseres Versuchsmateriales niiher zp erlautern, werden wir hier in miiglichster Kiirze die Rolle der Sauerstoffspannungen in dem respiratorischen Stoffwechsel unter- suchen. Ers tens ist es miiglich, dass diese Spannungen Bedeutung haben fur die Wanderung des Sauerstoffes zwischen Gewebe und Blut; entscheidend fur diese Wanderung oder nur bestimmend fur die Rich- tung derselben sind sie nicht,' es ist aber wohl anzunehmen, dass sie je den Umstanden nach unterstiitzend oder hemmend auf die Wan- dernng einwirken konnen. Ausserdem ist es wahrscheinlich, dass die Grijsse der Sauerstoffspannungen Einfluss auf die im Blute selbst vor sich gehenden Processe hat; indessen wissen wir hieriiber nichts Jlheres, ebensowenig wie wir irgend etwas Sicheres dariiber wissen, einen wie grossen oder kleinen Theil des sammtlichen respiratorischen Stoffwechsels die im Blute selbst stattfindenden chemischen Umsitze ausmachen. Unser Wissen iiber die Bedeutung der Spannungen in den zwei genannten Beziehungen ist dann nur gering.
Dagegen sind wir an einem anderen fiir den respiratorischen Stoffwechsel sehr wesentlichen Punkte im Stande, die Rolle der Span-
Bohr, Ueber die Lungenathmung. Dies Archiv. Bd. II.

UEBXR DEN SPECIFISCHEN SAUERSTOFFGEHALT DES BLUTES. 137
nungen niiher zu verfolgen; cs lasst sich nimlich nachweisen, dass diese entscheidcnd sind fur die proccntige Sauerstoffmenge in dem Plasma, welchcs dirccte Sauerstoffquelle fur die Gewebezelle ist. Dieses Verhiltniss, welchcs bisher, nach nieinem Ermcssen, nicht hinlanglich gewurdigt worden ist, bedarf einer ctwas niiheren Beleuchtung.
Das Plasma, wclches die Blutkiirperchcn umgiebt und cin un- umgiingliches Mittelglied ist fur die Gasbefirderung awischcn diesen und den Endothelzellen der Gefasse , enthilt selbst keine sauerstoff- bindenden, clissociablen Stoffe. Es absorbirt dcshalb den Sauerstoff im Wesentlichen auf dieselbe Weise, wio es das Wasser thut, also der Spannung proportionell und in geringer Menge. Dieses letzteren Um- standes wcgen hat das Plasma keinen nennenswerthen Vorrath an Sauer- stoff; in demselben Maasse, wie die Endothelzellen der Gefisse Sauerstoff vom Plasma hernehmen, muss dieser aus den in den Blutkijrperchen eingeschlosscnen Oxyhamoglobinen ersetzt werden. Der Ersatz geschieht so gut wie momentan wegen der gunstigen Diffusionsbedingungen zwischen dem Plasma und den Blutkorperchen , die eine besonders grosse Oberfliiche darbieten. In jedem Augenblick ist deshalb die Sauerstoffspannung in dem Plasma von der Sauerstoffspannung in den Blutkijrperchen abhiingig,’ da nun, wie oben angefuhrt, das Plasma sauerstofiindender Stoffe entbehrt, folgt hieraus, dass die Sauerstoff- menge im P la sma d i rec t mi t de r zu jeder Zeit i m Blu te h e r rsc h en d en S aue rs t offspan n u n g prop or ti on a1 i s t. Die Sauer- stoffmenge, die in jedem Augenblick den Zellen zu Gebote steht, ist dann direct und in einfachem Verhaltnisse von der Sauerstoffspannung im Blute abhangig, dagegen, und dieses ist wohl zu beherzigen, von der totalen Sauerstoffmenge des Blutes nur abhangig, insofern diese auf die Spannung Einfluss hat; in welchem Umfange und wie dieses stattfindet, werden wir im nachsten Abschnitte behandeln.
Die wichtige Rolle, welche auf diese Weise die Spannlingen fur die Sauerstoffzufuhr zu den Endothelzellen spielen, ist, wie man sieht, in dem Umstande begriindet, dass der snuerstoffbindende Stoff des Blutes in be- sonderen Behaltern, den Blutkorperchen, eingeschlossen ist ; in einer Losung von Hamoglobin wiirde das Verhdltniss ein ganz anderes sein. Die oben stehende game Betrachtungsweise lasst sich deshalb auch nicht einfach auf die KohlensKure im Blute uberfuhren, da sowohl im Plasma als in den Blutktirperchen kohlenshrebindende Stoffe vorhanden sind.
Es bleibt hiermit die Frage unbeantwortet, ob die Sauerstoffipannung im Plasma und den Blutkorperchen eine gleich grosse sei. Mit dieser Frage haben wir hier nichta zu thun, wie wir nur fur den Umstand Gebrauch haben, dass die Sauerstoffspannungen auf die angegebene Weise von einander ab- h8;ngig sind.

135 CHRISTIAN 12trrr1t:
111. Ueber das Verhiil tniss zwischen der totalen Sauers tof f - menge und dcr Blu tspannung des Blutcs.
So lange das Hiimogloljin nicht seine Modification verlindert, be- steht zu jeder Zeit ein bestimmtes Verhiiltniss zwischen dcr Sauerstoff- spannung des Blutes und dcr in demselhen cnthnltenen RIengc Sauer- stoffs; iibcr die Art dicses Vcrliiiltnisscs werden mir durch die Dis- sociationscurve des Hiimoglobins l)elehrt,l denn die Sauerstoffmenge des Blutes wird zum ganz uberwiegenden Theile von dcm in den Ulut- kurpcrchen an das HBmoglobin gebnndencn Sauerstoff ausgeniacht. Die Sauerstoffmenge dcs Plasmas ist dern gegeniiber ganz verschwin- dend. TVo man dagegen Vcrinderungcn in der Nodification des Hii- moglobins antrifft, hurt der fcste Zusammenhang zwisclien der Saucr- stoffmenge und der Sauerstoffspannung auf, wie es im erstcn Abschnitt dieses Capitcls dargestellt wurde.
In melchem Gradc und auf welche Weisc die Sanerstoffspannung von der Sauerstoffmenge abhiingig ist, ist eine Frage von grosser Wichtigkeit fur dic Rcspirationslehre, denn die Spannungcn, die zu joder Zeit der Saucrstoffmcnge in den das Plmna unmittelbar um- gebenden Endothelzellen proportional sind, wcrden dadurch das Maass fur das Sauerstoffangebot an die Zellen. Die Sauerstoffmengen im Blutc sind dagegen, menn dles Uebrige glcich, auf ,jedem Stadium des Blutlaufes durch die Capillaren eine Function des schon stattgehabten Sauerstoffverbrauches. Die Frage von dern Verhaltnisse zwischen Spannung und Sauerstoffmenge ist deshalb eine Frage von der Ab- hangigkeit des Sauerstoffangebotes yon dem schon stattgehabten Sauer- stoffverbrauch.
Betrachten wir zuerst den Fall, in welchem das Hiimoglobin wlh- rend des Kreislaufes seine Modification nicht verihdert. Wiihrend der stetigen Aufnahme und Abgabe von Sauerstoff, worin eine der Haupt- functionen des Blutes besteht, wird das Hiimoglobin bestandig seine Sauerstoffmenge wechseln, indem diese zwischen einem Maximum (im Arterienblute) und einem Minimum (im Venenblute) sich bewegt, wo- nach die Spannungen gesetzmassig steigen und fallen. Die neben- stehende Fig. 2 mag die Dissociationscurve des Hamoglobins dar- stellen; auf die genaue Form derselben kommt es hier nicht an, son- dern nur auf die Ziige, die fur die verschiedenen Hamoglobine bei rerschiedener Temperatur die gleichen sind. Es sind die folgenden:
Die Curve, deren Abscisse die Saucrstoffspannungcn, deren Ordinate die von 1 g Hiimoglobin aufgenommenen Sauerstolbengen sind.
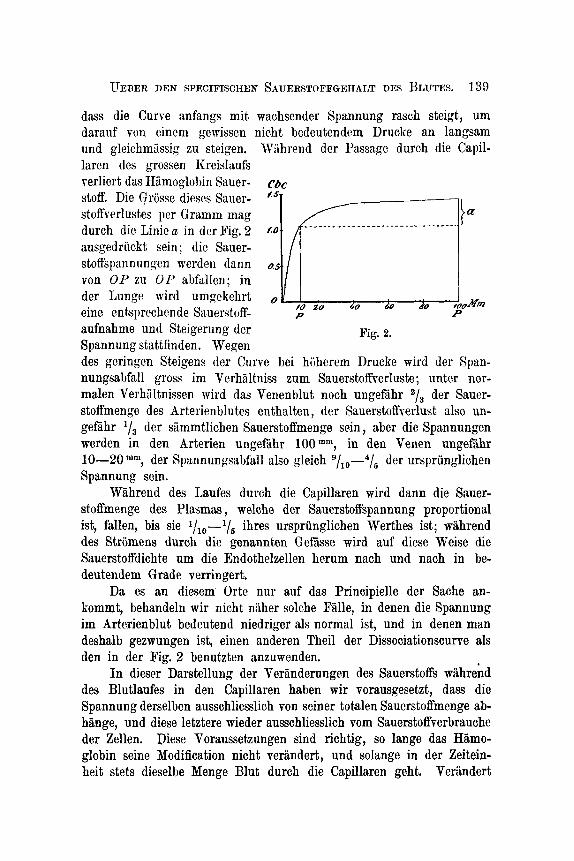
UEnER DEN SPECIFISCHEN SAUERSTOFPGEHALT DES BLUTES. 1 30
stoffverlustes per Gramm mag
ausgedriickt sein; die Sauer- stoffspannungen werdcn dann 05,.
von 01' zu OI' abfallen; in der Lunge wird umgekehrt
d"rch die Linic a in der Fig. ,,o - ------_--------------. l a

140 CHRISTIAN Llo~ilt:
dagegen &is Hiimoglobin seine Modification, hort, wie wir im 1 gesehen, der feste Zusammenhang zmischcn der Sauerstoffspannung und der Saucrstoffmenge auf, und insofern die in der Zciteinheit durch die Gefiisse gchcnde JIenge Blutes eine andere wird, ist, wie mir gleich naher sehcn werden, die Saucr\toffmenge nicht liinger ausschliesslich ahhiingig I om Sancirhtoffi-erIJri~uclie ron Scitcn tlcr %ellen. Es ist dann klar, dass dcm Organismus Xittel zu Gcbote stehen, uin die Sauerstoft- spannungen in den Capillaren innerhalb gewisser Grenzcn von dem dort stattfindenden Sauerstoffrerl~rauclic unahhlncig zu machcn, und dime Rlittel sind:
1) Ver i indernng der Dlenge des i n der Ze i te inhe i t d i e Cap i l l a r e n pass i re n d en H i m o glo hi n s und
2) eine withrend des l i re i s laufes s t a t t f i n d e n d e V e r a n - d e r u n g des specifischen Sauers tof fgehal tes des Blutes .
In Bezug auf das erstgenannte Regulirungsmittel ist cs einleuch- tend, dass je griisser die Mengc des in der Zeiteinheit die Capillaren passirenden Himoglobins ist, desto geringer ist, wenn der Sauerstoffver- branch als constant vorausgesetzt wird, der Bruchthcil des gcsammten Sauerstoffvcrlustes, der auf jedes Grainm Himoglobin fillt, desto geringer wird dann auch hierdurch der Spannungsabfall werden. Eine Ver- mehrung der Hiimoglobinmenge, welche in der Zeitcinheit ein Organ passirt, kann einestheils dadurch zu Stande kommen , dass der Reich- thum des Blutes an HHmoglobin griisser wird, theils dadurch, dass die Nenge des durohstriimenden Blutes vermehrt wird. Es ist nicht an- zunehmen , dass eine Zunahme des Himoglobins plotzlich stattfindet, dieselbe wird dagegen als eine hngsamere Art von Regulirung wirken kiinnen , wenn der Organismus allmihlich unter veriinderte Verhalt- nisse gebracht wird, z. B. durch Aufenthalt in hochliegenden Regionen.’ Die Menge des in der Zeiteinheit durch die Capillaren stromende Blut kann dagegen momentan wechseln, theils in einem einzelnen Organe durch Erweiterung der Blutgefisse desselben, theils was den ganzen Kreislauf betrifft , durch Vermehrung der Herzthatigkeit; in beiden Fallen resultirt hieraus eine Vermehrung der Menge des durch- stromenden Blutes, sodass, wenn alles ubrige gleich, also auch unter der Voraussetzung gleicher Beschaffenheit des Arterienblutes, die Be- dingungen fur die Sauerstoffaufnahme der Zellen gunstiger werden, indem die Sauerstoffmenge im Plasma weniger abnimmt als es sonst bei gleichem Sauerstoffverbrauche der Fall sein wurde. Es ist indess
Viault , Compt. rend. t. CXI. p. 917.

zu bemerken, dass, wo die Thatigkeit des Herzens vermehrt, dort auch der Kreislauf durch die Lungcn beschleunigt mird; es mird wohl dann miiglicher Weise die hijliere Geschmintligkei t eine meniger vollstliidige Slttigung des Blutes mit Sauerstoff mit sich fuliren kijnnen , wodurch die Sauerstoffspannung dcr Arterien fallen murde; hicrubcr l a s t sich nur durch Versuche in jedem Einzelfalle urtheilen.
Die gcnaiinte Regulirung mittels Veriinderungen in der Circulation wird selten fur sich allein mirken, denn wie wir schon im ersten Cn- pitel gesehen, geht normnler Weise eine s tc t ige Veri inderung des specifischen Snuerstoffgehnltes TOY sich, melche in den verschie- denen Gefissgebieten nicht dieselbe ist. Wenn, wie es haufig der Fall ist, der specifische Sauerstoffgehalt sich rerschieden im Arterien- und Venenblut zeigt und sich also miillrend des Laufes durch tlic Capillaren verandert, ist hiermit ein Mittel an die Hand gegeben, zur Verinderung der Sauerstoffspannung unabhangig von der totalen Sauerstoffmenge des Blutes also ein Mittel zur Regulirung der Sauerstoffmenge im Plasma, unabhangig von dem Sauerstoffverbrauche der Zellen. Und da ferner, wie wir es im ersten Abschnitt dieses Capitels gesehen haben, die Verinderung des spccifischen Sauerstoffgehaltes eine Wirkung der Gewebe auf das Blut ist, sind die Zellen durch dieses Mittel im Stande, selbs t einen regulirenden Einfluss auf die Sauerstofftlichte in dem die- selben unmittelbar umgebenden Medium auszuuben. Wegen des theil- weise geringen Steigens in dem als Regel im Organismus benutzten Stucke der Dissociationscurre (s. Fig. 2) kann ein ger inger Abfall des specifischen Sauerstoffgehaltes eine bed eu t e nd e Spannungsveranderung mit sich fiihren. Insofern der specifische Sauerstoffgehalt auf diese Weise in dem Beispiel, welches der Fig. 2 zu Grunde liegt, mit einem Werthe gleich der Linie a abfiele, wurde wiihrend des Capillardurch- laufes gar kein Spannungsabfall stattfinden; das Plasma wiirde mahrend des ganzen Laufes trotz des statthabenden Sauerstoffverbrauches seine procentige Sauerstoffmenge behalten.
[Es ist hier zu bemerken, dass die kleineren Verjnderungen des specifischen Sauerstoffgehaltes, die wir hiufig bei unseren Versuchen antreffen, keineswegs ausschliessen, dass die durch ihre Umbildung die Veranderung bewirkenden Hamoglobine rucksichtlich ihrer Sauerstoff- bindung in bedeutendem Grade verschieden sind, wie z. B. das y- und @-Hiimoglobines sind, denn da jedes einzelne Blutkorperchen fur die Veriinderungen ein besonderes Gebiet bildet, wird je nach der Zahl der beeinflussten Blutkorperchen jede Gradation des specifischen Sauer- stoffgehaltes vorkommen konnen.]

142 CIIRISTIAN BOHR :
R e s u m 6 e. Die Grijsse der Sauerstoffzufuhr zu den Zellen der Ge-
webe i s t von d e r Sauerstoffmenge im Plasnia und dicse v i e - derum von de r Snuerstoffspannung abhiingig. Die Sauer - s toffspannung is t i nne rha lb gewisser Grenzen von der totaler, Sauerstoffmenge des Blutes und dadurch von dem Snuerstoff- v er 1) rau c h c d e r Z e 11 en un a b h iin gig.
W. Einige Beispiele von Regul i rung mi t te l s Veri inderung d e s s p e ci f i s c h en 8 a u e r s t o f f g e h a 1 t e s.
Unsere Versuche sind zum grossten Theil auf das blosse Con- statiren der gesetzmiissigen Veriinderungen des specifischen Sauerstoff- gehaltes angelegt. In manchen Fallen haben die Versuche sich nicht weit genug erstreckt, um uns eine mehr detaillirte Einsicht in die Art der Regulirung zu verschaffen, besonders veil das Arterienblut allein untersucht morden ist, wahrend eben das Verhiiltniss zwischen dem Arterien- und dem Venenblute eine michtige Rolle bei der Re- gulirung spielt; ebenfalls hatte es Bedeutung gehabt, falls es miiglich gewesen , gleichzeitig die wirkliche Sauerstoffmenge im circuliren- den Arterien- und Venenblute, sowie die Grosse des respiratorischen Stoffwechsels zu bestimmen. Indem ich hoffe, bald Versuche vorlegen zu konnen, in denen diese sammtlichen Anspruche erfullt sind, be- schranke ich mich hier darauf, zwei Versuchsreihen zu discutiren, die schon jetzt im Wesentlichen mir hinlanglich beleuchtet erscheinen; ich habe dieselben hauptsachlich in der Absicht hier angefuhrt, urn sie
als Beispiele dienen zu lassen von der Anwendungsweise der in die- sem Capitel entwickelten Satze.
Ar te r ien- und Venenblu t (siehe erstes Capitel 11). Wahrend des Rlutlaufes durch die Musku- latur der hinteren Gliedmaassen wird normal der specifische Sauer- stoffgehalt rerringert, hierdurch wird dem Abfall der Spannung und der Sauerstoffmenge des Plasmas entgegengearbeitet , der wie oben
entwickelt eine Folge des Sauerstoffverbrauches der Gewebe ist. In Fig. 3 mag die Curve 1 die Dissociationscurve fur das Hamoglobin des Arterienblutes, die Curve 2 fur das des Venenbhtes bedeuten. Es mag
I
* P P Fig. 3.

UEBER DEN SPECIFISCHEN SAUERSTOFFGEHALT DES BLUTES. 143
ferner die Ordinate pa = PA die Sauerstoffmenge bedeuten, die nach dem Laufe durch die Capillaren per Gramm Hamoglobin ubrig ist; diese Sauer- stoffmenge wurde dann, insofern das Arterienblnt seinen specifischen Smerstoffgehalt behielte, den Druck p ausiiben. Wegen der in den Capillaren eingetretenen Veriinderung des specifischen Sauerstoffgehaltes ubt indess die gleiche Gasmenge den weit griisseren Druck P aus. Diese Spannungsregulirung wirkt innerhalb meiter Grenzen; normaler Weise ist der A bfall des specifischen Sauerstoffgehaltes vom Arterien- zum Venenblute bis des ganzen Werthes, bei der Curarevergiftung ist er aher verschwunden und bei der Pyocyaneusvergiftung bedeutend vermehrt bis zwischen lj4 und
Versu c h e mi t Ein a t hmun g sau ers t o ffar m er Lu f t (Versuche Nr. 40-43). Hier war vor dem Versuche der specifische Sauerstoff- gehalt im Arterienblute 406, im Venenblute 386; wihrend der Ein- athmung sauerstoffarmer Lift (8 Procent Sanerstoff) wurde die ge- nannte Grosse auf 388 im Arterienblute und auf 389 im Venenblute verandert. Es geschah also ein Abfall des specifischen Sauerstoff- gehaltes des Arterienblutes , wodurch dieses dem Veneublute gleich wurde. Um die Bedeutung dieser Veranderungen zu verstehen, ist noch.ferner zu bemerken, dass nach P. Ber t und anderen Forschern die Thatigkeit des Herzens wahrend des Athmens sauerstoffarmer Luft gesteigert wird, wahrend sich die in der Lunge in der Zeiteinheit auf- genommene Sauerstoffmenge etwas serringert. Haben wir das Recht, diesen Resultate auf unsere Versuchsthiere Anwendung zu geben, bei denen derartige Bestimmungen nicht gemacht wurden,l dann ist wiihrend des Athmens sauerstoffarmer Luft der Blutstrom durch die Lungen ein lebhafterer und in Verbindung hiermit die Sauerstoffauf- nahme eine geringere gewesen, welches zur Folge haben wird, dass jedes Gramm Hamoglobin eine geringere Snuerstoffmenge in sich auf- genommen hat. Hieraus wurde, wenn alles iibrige gleich, folgen, dass die Sauerstoffspannung im Arterienblute geringer wurde in Verbindung mit den hieran sich kniipfenden hinlanglich besprochenen Folgen ; indess wird diesem Spannungsabfalle in der That durch Verringerung des specifischen Sauerstoffgehaltes im Arterienblute entgegengewirkt Gleichzeitig ist der bevor der Einathmung von 8 Procent Sauerstoff sehr ausgesprochene Unterschied zwischen dem Arterien- und Venenblute ver- wischt worden; es versteht sicli, dass auch dieses eine Regulirung ist,
Dass diese zulbsig ist, liiest sich kaum bezweifeln; indess verdient'es doch ausdriicklich bemerkt zu werden, dass ich in meinem speciellen Versuche nicht die Thiitigkeit des Herzens und die Grssse des respiratorischen Stoff- wechsels untersucht habe.
des Werthes.

dean die eingetretene lebhaftere Circulation muss stets eine Be- gunstigung der Sauerstoffzufuhr zu den Zellen durch Vermehrung der Sauerst,offmenge im Plasma bewirken; das TOT den1 Einathmcn sauer- stoffarmer Luft vorhandene ebenfalls bep~nstigcnde r\loment, tler Lnter- schied des specifischen Saucrstoffgehaltes zwischen dem ilrterien- unrl dem Venenblut ist dann fortgefdlen.
Die gefundeneii VeAnderungen in Blut und Circulation zeigen sich dann als eine Hegulirung, welche den Zweck hat, die Sauerstoff- zufuhr zu den Zellen mijglichst unabhiingig z u machen von der statt- gehabten Verlnderung in der Zusammensetzung der Einathmungsluft;.
Zum Schluss miichte ich nocli aufmerksam machen auf die enge Verbindung, welche bei der in dieser Abhandlung beschriebenen Re- gulirung der Gemeberespiration zwischen s5mmtlichen Organen des Korpers zu Tage tritt. Der specifische Sauerstoffgehalt, den die ver- schiedenen Organe in dem sie durchstrijmenden Blute verandert haben, ist wahrend jedes Kreislaufes im Arterienblute wieder auf seinen ur- sprunglichcn Werth zuruckgebracht worden. Ob diese letztere Ver- anderung durch das in den oberen Theil der Vena cava einstromende Blut oder in den Lungen geschieht, ist noch nicht untersucht worden, jedenfalls kann die Wirkung der Regulirunq nur durch eine bestan- dige genau abgewogene Wechselwirkung verschiedener Organe zu Stande kommen.

![Filmfestival „ueber Mut“ · Filmfestival „ueber Mut“ [Donnerstag, 25.11. bis Mittwoch, 01.12.2010] Das Festival „ueber Mut“ der Aktion Mensch zeigt zehn internationale](https://img.pdfslide.org/doc/110x75/5e42d977b10a0a701f0f1034/filmfestival-aueber-mutaoe-filmfestival-aueber-mutaoe-donnerstag-2511-bis.jpg)