Embed Size (px)
Citation preview

Acta histochem. Bd. 57, S. 212 - 234 (1976)
Ana.tomisches Institut del' Ernst·Moritz·Arndt·Universitat Gl'eifswald
(Direktor: Prof. Dr. sc. med. R. SULZMANN)
Zur Enzymhistochemie des Pankreas del' Sandl'atte (Psammomys obes'us) nach calol'isch diffel'enter Ernahrung
Enzyme histochemistry of the sand rat pancreas (Psammomys obesus) after different caloric food
Von ALFRED DORN und GERDA KOCH
Mit 17 Abbildungen
(Eingegangen am 15. November 1975)
Zusammenfassung
1m Rahmen einer Komplexuntersuchung werden 12 ausgewiihlte Enzyme (Oxidoreductasen und
Hydrolasen) am Pankreas normoglykiimischer Sandratt~n (Psammomys obe8us) histochemisch na.chgewiesen . Die Sandratte neigt zur spontandiabetischen Stoffwechsellage, wenn sie im Vivarium ge
halten wird. Das Material stammte von Tiergl'uppen , die ad libitum bzw. nach einem bestimmten
Diatprogramm erniihrt wurden. Das topochemische Enzymmnster in Langerhansschen Inseln und exokrinen Acini konnte zwischen den einzelnen experimentellen Tiergruppen und mit dem gut untersuchten Modell del' Wistar·Ratte verglichen werden. Da enzymhistochemische Untersuchungen am Sandrattenpanlu eas bisher fehlten, wurden die Ergebnisse den spiirlichen einschlagigen Befunden anderer Tiere und des Menschen gegeniibergestellt und im Zusammenhang mit der veranderten Stoffwechsellage unter besonderen Erniihrungssituationen und beim Diabetes diskutiert.
Summary
In a complex investigation were evidenced 12 selected enzymes (oxidoreductases and hydrolases)
in the pancreas of normoglycemic sand rats (Psammomys obes1ts). The sand rat developed a spontane
ous diabetic metabolism by holding in the vivarium. The material based of animal groups that were
fooded ad libitum or after defined dietetic regimes. The topochemicaI pattern of enzymes in the
LANGERHANS' islets and exocrine a cini can be compared between the sand rat groups and with the
good investigated model of the Wistar rat. Because enzymhistochemical investigations of the sand
rat pancreas absented, were the results compared wit,h the rare specific observations of other animals
and the man and the changed metabolic level were discussed under the especially food situation and
in diabetes mellitus.
Seit einiger Zeit wird die in den Wustengebieten des Orients und Nordafrikas beheimatete Sandratte (Psammomys obesus) in del' experimentellen Diabetesforschung als Versuchstier eingesetzt, da sie unter labormaBigen Haltungsbedingungell zum spontanen Diabetes mellitus neigt und damit am ehesten naturliche Diabetesformen imitiert . Die Zucht ist im Vivarium auBerordentlich problematisch (BRUNN 1971, HAHN et aI. 1971). Es ist nicht uninteressant zu wissen, daB der spontane Diabetes

Zul' Enzymhistochemie des Pankl'eas del' Sandl'atte 213
bei der Sandratte offenbar durch die veranderte Ernahrungssituation ausge16st wird, also eine Art Zivilisationskrankheit darstellt. Untersuchungen an der Sandratte als spontandiabetischem Tiermodell erscheinen besonders interessaIlt, zumal uns keine histotopochemischen Untersuchungen am Pankreas dieser Tiere bekannt geworden sind. Der Einsatz dieser Tiere fUr die medizinisch-biologische Forschung wird dadurch erschwert, daB hohe Absterberaten wahrend des Transportes auftreten und auch wahrend der Adaptationszeit nicht ausbleiben. Wurden die Tiere in der Gefangenschaft mit herkommlichen LaborpreBlingen gefUttert, so zeigten die Sandratten Gewichtszunahme, Hyperglykamie und Glukosurie bei verminderter Glukosetoleranz und Hyperinsulinamie mit einer im Verlauf zunehmenden Degranulation und Vakuolenbildung in den B-Zellen. Erfolgt eine Ernahrung mit frischem Gemuse, so treten diese Veranderungen im allgemeinen nicht ein (MIKr et al. 1967). Durch calorische Reduktion (30 Kcal(d) bei vorher ad libitum mit Ruben ernahrten Sandratten trat innerhalb von 4 bis 6 W ochen eine N ormalisierung der Glukosetoleranz, eine Senkung des Korpergewichtes und des lnsulinspiegels ein. Eine Zufuhr von Mineralien und Vitaminen verbessert die Vitalitat der Tiere (PRANGE 1968).
Wir haben uns bei unseren Untersuchungen ausschlieBlich mit dem histotop:)chemischen Enzymmuster des Sandrattenpankreas nach unterschiedlichen Diiitregimes beschiiftigt. Durch die zur VerfUgung stehende Materialmenge waren wir gezwungen, die Palette der uns interessierenqen Enzyme einzuschranken und eine sorgfaltige Auswahl der Methoden zu treffen.
Da uns histochemische Untersuchungen am Pankreas der Sandratte Dicht bekannt sind, haben wir in erster Linie solche Enzyme untersucht, die charakteristisch fUr die Langerhanssche lnsel sind und als histochemische Leitenzyme fUr die Langerhanssche lnsel aufgefaBt werden konnen (G6PDH, sPase, lCD-NADP). Wir erweiterten unser Programm urn weitere 9 Enzyme, die uns im Zusammenhang mit dem biochemischen Geschehen interessant erschienen.
Material und Methode
Die normale Ernahl'ung del' Sandl'atte besteht aus Pflanzen, die einen l'elativ hohen Salzgehalt besitzen.
Unsel'e Vel'suchsol'dnung:
Basisgl'uppe
< 30 Kcaljd 14 bis 21 d
< 30 Kcaljd 28 bis 35 d
> 40 Kcaljd 14 bis 21 d
> 40 Kcaljd 28 bis 35 d
Die Nahrung fUr die Basisgruppe bestand aus Griinkohl und Chinakohl. Die Zusammensetzung
del' Nahrung del' andel'en Gl'uppen war wie folgt:
30 Kcal: 83 % Wasser, 8,3 % Kohlenhydrate, 1,7 % Fett, 5,1 % EiweiB, 1,4 % Asche
40 Kcal: 66,5 % Wassel', 14,5 % Kohlenhydrate, 7,3 % Fett, 9,9 % EiweiB und 2,1 % Asche.
Die Differenz zwischen beiden Diatgruppen betrug bei del' 40 Kcal.Nahrung 20 % Wasser
weniger, 75 % Kohlenhydrate mehr, 322 % Fett mehr, 94 % EiweiB mehr und del' Ascheanteil wurde um 48 % erh6ht. Parallel dazu wurden weiBe Wistarratten (Rattu8 norvegicu8 B) untersucht, die in

214 A. DORN und G. KOCH
der Ernahrungslage der "Basisgruppe" der Sandratten entsprachon. Teile des milznahen Pankreas wurden unfixiert tiefgefroren und im Kryostaten bei --18°C in Schnitte von 10 (.Lm Dicke zerlegt. Strukturerhaltung sowie die Erhaltung der Enzymaktivitat im Gewebe waren gut. Histochemisch
wurden folgende Enzyme nachgewiesen:
Enzym
1. Oxidoredu6tasen
1.1. Glukose-6-Phosphat-Dehydrogenase
1.2. Lactatdehydrogenase 1.3. p-Hydroxibutyratdehydrogenase
104. NAD-abhangige Isocitratdehydrogenase 1.5. NADP-abhangige Isocitratdehydrogenase
1.6. Succinatdehydrogenase
1. 7. NAD-abhangige Malatdehydrogenase
1.8. NADP-abhangige Malatdehydrogenase
2. Hydrola8en
2.1. Glukose-6-phospahtase
2.2. saure Phosphatase 2.3. Adenosintriphosphatase (pH = 9,4)
204. Adenosintriphosphatase (pH = 7,2)
Abkiirzung
G6PDH
LDH HBDH
ICDHjNAD
ICDHjNADP SDH
MDHjNAD
l\IDHjNADI'
G-6-Pase s Pase
a ATPase (n. PADYKULA und HERMAN)
n ATPase (n. WACHSTEIN und MEISEL)
EC. Nr.
1.1.1.49
1.1.1.27
1.1.1.30 1.1.1.41 1.1.1.42 1.3.99.l.
1.1.1.38
1.11.40
3.1.3.9.
3.1.:1.2.
3.6.l.3.
3.6.1.3.
Die Medien fiir die histochomischen Enzymnachweise wurden nach Angaben von folgenden Autoren zusammengestellt: PEARSE (1972), ARNOLD (1966), GEYER (1969).
Photographische Technik: NfpK, Miflex-Standard, Belichtungsautomatik, Planapochromate II), und 16, Planachromate 40 und HJ 100, Projektiv 3,2 (alles VEB Carl-Zeiss-Jena).
Filmmaterial: ORWO NP 15, Vephota-Papiere. Neben den enzymhistochemischen wurden parallel dazu biochemische und histologische Unter
suchungen von anderen Arbeitsgruppen durchgefiihrt (KOHLER et al. 1975, HAHN VON DORSCHE',
et al. 1975).
Befunde
Allgemeine Vorbetrachtungen
Obwohl die Sandratte in der experimenteIlen Diabetesforschung schon seit vielen Jahren eingefiihrt ist, gibt es erstaunlich wenig Ver6ffentlichungen iiber morphologische Untersuchungen an Langerhansschen Inseln dieses wichtigen, aber schwer
ziichtbaren Tieres. Enzymhistochemische Untersuchungen sind uns bisher nicht bekannt geworden. Wir glauben, unserem spezieIlen Versuchstier die vielfach untersuchte adulte Wistarratte gegeniibersteIlen zu k6nnen. Bei einem Vergleich der beiden Spezies £aIlt jedoch ein unterschiedliches Enzymmuster auf. Die Basisgruppe unserer Experimente war aus einem homogenen Tiermaterial zusammengesetzt, bei dem die Stoffwechselparameter biochemisch als relativ einheitlich registriert werden konnte. Die vier weiteren Tiergruppen wurden den experimenteIlen Beeinflussungen des. Stoffwechsels durch hypocalorische bzw. hypercalorische Ernahrung unterworfen.

Zur Enzymhistochemie des Pankreas der Sandratte 215~
In der nachfolgenden Tabelle sind unsere Untersuchungsergebnisse zusammengefa13t. Hier k6nnen wir das differenzierte Verhalten der Enzymaktivitiiten in den. inselspezifischen Zellen im Vergleich zu exokrinen Pankreaszellen als auch die relativen Enzymaktivitaten bei den verschiedenen Versuchsgruppen der Sandratte und der adulten, normalen Wistarratte ablesen.
Der besseren Ubersicht wegen werden wir die Befunde der einzelnen Tiergruppe~ getrennt darstellen und in besonderen Fallen gegeneinander vergleichen.
I~ Wistarrafle 8asisgruppB <30ffw//d
!f.-21d
Enzym ABE A B E A B E
(}5 PDH t Ht! >0 tt tt t t t t
LDH + + itt tt ttitt tt tt ttt i---
tt!tt HBDH + t (t) tt + + + JCDH NAD + t (t) + t + t + + JCDH NADP + tt + ++ tt 1+ tt ++ + SDH (t) ++ + + t(t) t + + +
. I1DH NAD t ttt ttt t t t tt tt ttt
NDH NADP + tt + + Ht) + + + + ;; Pase 0 H+ 0 tH+ 0 0 H+ 0 (}-5-Pase 0 0 0 0 0 0 0 0 .0
a ATPase tt tt ++ 0 0 0 0 0 0
j n ATPase 0 0 0 ++ tt + + t +
1. Basisgruppe
1.1. Oxidoreductasen
1.1.1. G lukose- 6 -phospha tdehydrogenase
G6PDH, EC 1.1.49
(JOffca/ /d >'to/{ca! /d >~Of(ca//d 28-3Sd 1ft-21d 28-35d
A B E A B E A B E
t t t + tt t !
tt tt tt tt +t Ht t t t tt tt t+ H+) Ht) tt<t) +tltt tt
+ + + t + + +1+ t
++ ++ + tt tt + ++1++ + ++ tH ++ ++ Ht tt tt H+ tt
++ + tt tit t ++ tt + + ++ + + + + + + + 0 0 0 0 tt 0 0 0 0
0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0
++ tt + + + + ++ ++ t
In den A- und B-Zellen der Langerhansschen Inseln lie13 sich dieses Ferment gut·, nachweisen. Dadurch hoben sich die Inselzellen deutlich von den schwach reagieren-. den Acinuszellen abo
1.1.2. Lactatdehydrogenase
LDH, EC 1.1.1.27. Der Nachweis der LDH lie13 in dieser Tiergruppe keine topochemische Differen
zierullg der Zellen zu, da sowohl die endokrinen als auch die exokrinen Zellen eine. gleich gute Aktivitat erkennen lassen. Die Inseln mu13ten nach rein morphologi.schen.. Gesichtspunkten gefundell werden.
1.1.3. fJ-Hydroxi butyratdehydrogenase
HBDH, EC 1.1.1.30.

216 A. DORN und G. KOCH
Almlich dem Nachweis der LDH gestattete die gute, aber flir alle Zellen des Pankreas gleichmiLBige Aktivitiit keine topochemische Differenzierung der Inseln.
1.1.4. Isocitratdehydrogenase
ICDH, NAD-abhiingig, EC 1.1.1.41. ICDH, NADP-abhiingig, EC 1.1.1.42. Die NAD-abhangige ICDH bestach hier durch eine sehr schwache uniforme Enzym
reaktion in allen drei Zelltypen. AuffiHlig war eine ungewohnlich grobkornige Pd.zipitation des Diformazans in den Inselzellen. Hingegen lieB die NADP-abhiingige ICDH eine geringe Zelldifferenzierung zu. Die grobkornige Reaktion in den A- und B-Zellen war starker ausgepriigt als die feingranuliire in den Acinuszellen.
1.1.5. Succinatdehydrogenase
SDH, EC 1.3.99.l. Der Nachweis del' SDH fiel im gesamten Pankreas schwach aus und war durch
Gleichformigkeit gekennzeichnet, die wir in der 1. Basisgruppe nicht in diesem MaBe beobachten konnten. Die B-Zellen zeigten lediglich im unmittelbaren perinukleiiren Zytoplasma eine geringfiigig verstarkte Reaktion dieses ubiquitaren Enzyms der Mito
chodrien.
1.1.6. Malatdehydrogenase
MDH, NPD-abhiingig, EC 1.1.1.38. MDH, NADP-abhiingig, EC 1.1.1.40. Auch dieses Ferment des Krebszyklus zeigte gegeniiber dem topochemischen Ver
halten der 1. Basisgruppe nur geringfiigige Differenzen hinsichtlich der Reaktionsstarke der untersuchten 3 Zelltypen. Auffallend war sowohl bei der NAD- als auch bei del' NADP-abhangigen Malatdehydrogenase die grobkornige Ablagerung des Diformazans. Hier gab es die schon oben erwiihnten krassen Unterschiede zur Wistarratte.
1.2. Hydrolasen
1.2.1. saure Phosphatase
sPase, EC 3.1.3.2. Die histochemische Verteilung dieses Enzyms war typisch und entsprach etwa
der der Wistarratte. Auffallend war die starke zytoplasmatische Reaktion in den B-Zellen und ein sehr schwacher Reaktionsausfall in den A-Zellen. Acinuszellen blieben absolut negativ.
1.2.2. Glukose-6-phosphatase
G6Pase, EC 3.1.3.9. Das wichtige Schliisselferment der Glykolyse konnte histochemisch nicht nach
gewiesen werden.

Zur Enzymhistochemio des Pankr'eas del' ~andl'atte
1.2.3. Adenosintriphosphatase
aATPase pH = 9,0, EC 3.6.1.3.
2L7
Dieses Ca ++ -abhiingige energieliefernde Ferment konnte ebenfalls nicht nachgewiesen werden.
1.2.4. neutrale Adenosintriphosphatase
nATPase, pH = 7,2, EC 3.6.1.3. Die Mg++ -abhiingige ATPase konnte durch eine gute Reaktion in den A- und B·
Zellen nachgewiesen werden. Wesentlich schwiicher stellten sich die Acinuszellen dar. Eine starke ATP-ase-ReaktionlieB die GefiiBwand in den Pankreasschnitten erkenncn.
2. Erniihrungsgruppe: <30 Kcaljd
Dauer des Experimentes: 14 bis 21 Tage
2.1. Oxidoreductasen
2.1.1. G lukose- 6 -phos pha tde hydrogenase
G6PDH, EC 1.1.1.49. Es iiberraschte eine eindeutige Abschwiichung der normalerweise hervorstechen
den B-Zellreaktion. In allen drei untersuchten Pankreaszelltypen fanden wir nunmehr eine schwache Reaktion des Enzyms des Pentosephosphatshunts.
2.1.2. Lactatdehydrogenase
LDH, EC 1.1.1.27. Die endokrinen Inselzellen reagierten gleichmiiBig und gut. Hingegen zeigten die
exokrinen Pankreaszellen eine sehr starke Aktivitiit, die gegentiber der Basisgruppe hervorstach.
2.1.3. {J -Hydroxi bu tyra tdehydrogenase
HBDH, EC 1.1.1.30. Dieses Ferment konnte in den untersuchten 3 Zelltypen des Pankreas ohne wesent
liche Unterschiede nur schwach llachgewiesen werden.
2.1.4. Isocitratdehydrogenase
ICDH, NAD-abhiingig, EC 1.1.1.4l. ICDH, NADP-abhiingig, EC 1.1.1.42. Die NAD-abhiingige ICDR zeigte eine schwache gleichmiiBige Reaktiou in den
Zellen des enda- und exokrinen Pankreas. Hingegen fanden wir in den A- und BZellen der Langerhansschen Inseln eine deutlich verstiirkte Aktivitiit der NADP
abhiingigen ICDR.
2.1.5. Succinatdehydrogenase
SDH, EC 1.3.99.1. Die SDH-Reaktion fiel in allen !3 Zelltypen des Pankreas gleichmiiBig schwaeh
aus, so daB dcutliche Unterschiede zur Basisgruppe und zur Enzymreaktion bei dee Wistarratte nachgewiesen werden konnten.
15 Acta histochem. Bd. 57

218 A. DORN und G. KOCH
2.1.6. Malatdehydrogenase
MDH, NAD-abhangig, EO 1.1.1.38. MDH, NADP-abhangig, EO 1.1.1.40. Die NAD-abhiingige MDH-Reaktion verlief in den Pankreaszellen recht stark.
Eine besonders hohe Aktivitiit zeigten die Acinuszellen. Demgegeniiber fiel die Reaktion der NADP-abhangigen MDH in den A-, B- und Acinuszellen gleichmiiBig und sehr schwach aus. Beide Fermente lieBen demnach Unterschiede zur Basisgruppe erkennen.
2.2. Hydrolasen
2.2.1. saure Phosphatase
sPase, EO 3.1.3.2. Wie bei der Wistarratte erschienen hier die B-Zellen der Langerhansschen lnseln
sehr aktiv. A- und Acinuszellen reagierten nicht. Die saure Phosphatase konnte auch bei dieser Erniihrungsgruppe als topochemisches Leitenzym fUr die Langerhansschen lnseln betrachtet werden.
2.2.2. Glukose-6-phosphatase
G6Pase, EO 3.1.3.9. Dieser Nachweis verlief im gesamten Pankreas negativ.
2.2.3. alkalische Adenosintriphosphatase
aATPase, pH = 9,0 EO 3.6.1.3. Die Oa++-abhiingige ATPase konnte ebenfalls nicht nachgewiesen werden.
2.2.4. neutrale Adenosintriphosphatase
nATPas3, pH = 7,2 EO 3.6.1.3. Die Mg++ -abhiingige ATPase zeigte gegeniiber der Basisgruppe 2 eme abge
schwiichte Reaktion in den Zellen der Langerhansschen lnseln, so daB aIle untersuchten Zellen des Pankreas eine uniforme, sehr schwache Aktivitiit zeigten.
3. Erniihrungsgruppe: < 30 Kealld
Dauer des Experimentes: 28 bis 35 Ta,qe.
3.1. Oxidoreductasen
3.1.1. G 1 ukose -6 -phospha tdehydrogenase
G6PDH, EO 1.1.1.49. Das Fermentmuster der G6PDH war bei dieser Tiergruppe durch eine schwache,
fast uniforme Reaktion in den A-, B- und Acinuszellen gekennzeichnet. Gelegentlich wurden die B-Zellen schwach hervorgehoben. Die Langerhanssche lusel muBte hier auf Grund von morphologischen Kriterien beschrieben werden.
3.1.2. Lactatdehydrogenase LDH, EC 1.1.1.27.

Zur Enzymhistochcmie des Pankreas del' Sandratte 219
Eine deutliche, gleichformige Reaktion war allen drei untersuchten Zelltypen eigentiimlich.
3.1.3 . P -Hydroxi bu tyra tdehydrogenase
HBDH, EC 1.1.1.30. Dieses Ferment zeigte ebenfalls eine gleichformige Zellvert eilung III den unter
suchten Zelltypen, abel' es war eine deutliche Aktivitiitssteigerung gegeniiber der zuvor beschriebenen Tiergruppe zu beobachten. Diese Aktivitiitsintensitiit wiire der Basisgruppe 2 vergleichbar.
3.1.4. Isocitl'atdehydrogenase
ICDH, NAD-abhiingig EC 1.1.1.41. ICDH, NADP-abhiingig EC 1.1.1.42. Die NAD-abhiingige ICDH zeigte erneut eine schwache, unifol'me Reaktion in
den P ankreaszellen. Eine deutliche Reaktionssteigerung lieBen, wie schon bei del' Basisgruppe 2 und del' zuvor besprochenen Tiergruppe, die A- und B-Zellen beim Nachweis del' NADP-abhiingigen ICDH erkennen.
3.1.5. Succinatdehydrogenase
SDH, EC 1.3.99.1. Eine deutliche Aktivitiitssteigerung bei diesel' Tiergruppe erfiihrt die SDH. Die
A- und Acinuszellen lassen eine gute Aktivitiit erkennen, die jedoch an Intensitiit von den B-Zellen iibertroffen wurde. Dieses mitochondriale Atemferment scheint beziiglich del' Erniihrungssituation sehr empfindlich zu sein.
3.1.6. Malatdehydrogenas e
MDH, NAD-abhangig, EC 1.1.1.38. MDH, NADP-abhiingig, EC 1.1.1.40. Gegeniiber den zuvor beschriebenen Untersuchungsserien ergaben sich hier geringe
Unterschiede im Enzymmustel' . Die NAD-abhiingige MDH lieB eine bemerkenswerte Abschwiichung del' Reaktion in den B-Zellen erkennen, bei gutel' Aktivitiit in den A- und Acinuszellen. Bei der N ADP -abhiingigen MDH war das Reaktionsverhiiltnis umgekehrt. Die B-Zellen zeigten gegeniiber den A- und Acinuszellen eine verstiirkte R eaktion.
3.2. Hydl'olasen
3.2.1. saure Phosphatas e
sPase, EC 3.1.3.2. Den sicher iiberraschendsten Befund lieferte die sPase. Gegeniiber allen bisher
untersuchten Gruppen erschienen alle Zellen des Pankreas l'eaktionslos. Es reagierten im sichel' inselhaltigen Pankreasschnitt auch nicht die B-Zellen. Uberkreuzversuche mit dem gleichen Inkubationsmedium lieBen keinen Zweifel an del' einwandfreien Methodik. Diesel' Befund wird besonders zu diskutieren sein.
15*

220 A. DORN und G. K00H
3.2.2. Glukose-6-phosphatase
G6Pase, EC 3.1.3.9. 1m gesamten Pankreas erhielten wir einen negativen Reaktionsausfall.
3.2.3. alkalische Adenosintriphosphatase
aATPase Ph = 9,0 EC 3.6.1.3. Ebenfalls negativer Reaktionsausfall im gesamten Pankreas.
3.2.4. neutrale Adenosintriphosphatase
nATPase pH = 7,2 EC 3.6.1.3. Die A-Zellen lieBen eine gute, die Acinuszellen eine schwache Reaktion del' Mg~~ ·
-abhangigen ATPase erkennen.
4. Ernahrungsgruppe: > 40 Kealld Dauer des Experimentes: 14 bis 21 Tage.
4.1. Oxidoreductasen
4.1.1. G 1 ukose- 6-phospha tde hydrogenase
G6PDH, EC 1.1.1.49. Wie bei den Tieren der B 'l-sisgruppen erschienen hier die B-Zellen besonders G6PDH.
aktiv. Dadurch hoben sich die Inseln sehr deutlich yom iibrigen sehr schwach reagierenden P ankreas abo
4.1.2. Lactatdehydrogenas e
LDH, EC 1.1.1.27. Dieses Ferm3nt re:t.sierte auch bei diesel' Tiergruppe sehr intemiv. Besonders die
Acinuszellen fielen durch eine hohe Aktivitiit auf, so daB die Inseln als gut reagierende, aber gegeniiber dem exokrinen Pankreas hell erscheinende Bezirke sichtbar werden.
4.1.3 . P -Hydroxybutyratd ehydrogenase
HBDH, EC 1.1.1.30. Die HBDH reagierte ahnlich del' LDH in den Pankreaszellen diesel' Tiergruppe
unterschiedlich. In den helIeren Inselbezirken sind A- und B-Zellen gut aktiv. Die exokrinen Pankreaszellen reagierten jedoch stark und gaben einen Kontrast zu den Inseln, die dadurch gut auffindbar waren.
4.1.4. Isocitratdehydrogenase
!CDH, NAD-abhiingig EC 1.1.1.41. !CDH, NADP-abhangig EC 1.1.1.42. Gegeniiber den zuvor untersuchten Sandrattenkollektiven gab es auch hier keine
nennenswerten Unterschiede. Bei del' NAD-abhangigen ICDH reagierten aIle untersuchten Zellen gleich schwach. Die Aktivitat del' NADP-abhiingigen !CDH war in den A- und B-Zellen starker als in den AcinuszelIen.
4.1.5. Succinatdehydrogenase
SDH. EC 1.3.99.1.

Zur Enzymhistochemie des Pankreas del' Sandratte 221
Die SDH war im Pankreas dieser Dntersuchungsgruppe gut nachweisbar. Die starkste Aktivitat zeigten die B-Zellen, so daB die rnseln sich optisch gut vom exokrinen Pankreasanteil abhoben.
4.1.6. Malatdehydrogenas e
MDH, NAD-abhangig EO 1.1.1 .38. MDH, NADP-abhangig EO 1.1.1.40. Die Reaktionen der beiden Malatdehydrogenasen uberraschten sehr, da sowohl das
NAD- als auch das NADP-abhangige Enzym in den A- und B-Zellen der Langerhansschen rnseln und in den Acinuszellen nur sehr schwach reagierte. Es erschien ein sehr gleichmaBiges Pankreasbild.
4.2. Hydrolasen
4.2.1. saure Phosphatase
sPase, EO 3.1.3.3.2. Dieses topochemische Leitenzym fUr die rnseln des Pankreas erfUllte bei dieser
Tiergruppe seine volle Funktion. Die B-Zellen reagierten gut bis sehr gut, aIle ubrigen Pankreaszellen (A- und Acinuszellen) waren inaktiv, d . h. in ihrem Zytoplasma war keine saure Phosphatase nachweisbar.
4.2.2. Glukose-6-phosphatase
G6Pase, EO 3.1.3.9. 1m gesamten Pankreasmaterial ein negativer Reaktionsausfall.
4.2.3. alkalische Adenosintriphosphatase
aATPase, pH = 9,0 EO 3.6.1.3. Ebenfalls negativer Reaktionsausfall im gesamten P ankreas.
4.2.4. neutrale Adenosintriphosphatase
nATPase, pH = 7,2 EC 3.6.1.3. Wir fanden in den A-, E- und Acinuszellen eine gegeniiber den anderen Dnter
suchungsgruppen abgeschwachte gleichmaBige Reaktion der Mg++ -abhangigen ATPase.
5. Ernahrungsgruppe: > 40 Kealld
Dauer des Experimentes: 28 bi8 35 Tage.
5.1. Oxidoreductasen
5.1.1. Glukose-6 -phosphatdehydrogenase
G6PDH, EO 1.1.1.49. Aus technischen Grunden konnte der Nachweis nicht gefUhrt werden.
5.1.2. Lactatdehydrogenase
LDH, EO 1.1.1.27.

222 A. DORN und G. KOCH
Eine uberraschend abgeschwiichte abel' gleichmiiBige Reaktion kennzeichnete die LDH-Aktivitiit.
5.1.3. tJ- Hydroxybutyra tdehydrogenase
HBDH, EO 1.1.1.30. Dieses Ferment zeigte eine gut ausgepragte indifferente Reaktion im Pankreas
schnitt. Die Zelltypen waren topochemisch nicht differenzierbar.
5.1.4. Isocitratdehydrogenase
ICDH, NAD-abhiingig, EO 1.1.1.41. ICDH, NADP-abhiingig, EO 1.1.1.42. Auch bei dem letzten Tierkollektiv gab es keine bemerkellswerten Unterschiede
hinsichtlich del' Aktivitiitsstarke und del' Zelldifferenzierung beim Nachweis del' bei
den Isocitratdehydrogenasen gegenuber den bisher beschriebellen Gruppen. Del' uniformen und schwachen Reaktion der NAD-abhiingigen IODH steht eine starke Reaktion in den A- und B-Zellen del' NADP-abhangigen ICDH gegenuber.
5.1.5. Sue cina tdehydrogenase
SDH, EO 1.3.99.1. Die SDH reagierte wie bei den meisten schon beschriebenen experimentellen Tier
gruppen besonders stark in den B-Zellen und gut in den A- und Acinuszellen. Die Diformazanniederschliige waren in der Regel recht grob.
5.1.6. Malatdehydrogenase
MDH, NAD-abhiingig, EO 1.1.1.38. MDH, NADP-abhangig, EO 1.1.1.40. Beim Nachweis diesel' beiden Enzyme gab es eine Reihe von Differenzen hinsicht
lich des Verteilungsmusters. Die NAD-abhiingige MDH reagierte in den A- und BZellen stiirker als in den Acinuszellen. Die NADP-abhiingige MDH I'eagierte sclnvach bis gut in den 3 untersuchtell Zelltypen des Pankreas.
5.2. Hydrolasen
5.2.1. saure Phosphatase
sPase, EO 3.1.3.2. In diesel' Untersuchungsserie fiel del' topochemische Nachweis des wichtigen hydro
lytischen Enzyms negativ aus. Ebenso wie bei del' hypokalorisch langzeitig erniihrten Sandratte zeigten die B-Zellen keine Aktivitiit del' sauren Phosphatase.
5.2.2. Glukose-6-phosphatase
G6Pase, EO 3.1.3.9. NegativeI' Reaktionsausfall im gesamten Pankreasmaterial dieses Tierkollektivs.
5.2.3. alkalische Adenosintriphosphatase
a ATPase , pH = 9,0, EO 3.6.1.3. Ebenfalls negativer Reaktionsausfall.

Zur Enzymhistochemie des Pankreas del' Sandratte
6.2.4. neutrale Adenosintriphosphatase
nATPase, pH = 7,2, EC 3.6.1.3.
223
Wie schon mehrfach in den Gruppen beobachtet, war die Aktivitat der Mg++ -abhiingigen ATPase in den A- und B-Zellen starker ausgepragt als in den exokrinen Pankreaszellen. Die Reaktion dieses Fermentes in den GefaIlen war durchaus bemerkenswert, stellt aber in unser en Untersuchungen nur einen Nebenbefund dar.
Diskussion
D el' Histochemie del' Langerhansschen Insel des Pankreas ist in den letzten Jahren viel Aufmerk
samkeit gesch enkt worden, denn die hormonale Aktivitat des endokrinen Pankreas und dic Klarung der funktionellen Bedeutung der einzelnen Zelltypcn sind mit histochemischen Problemen eng vel'·
knupft . Inzwischen ist sogar bel'eits mehrfach del' indirekte immunohistochemische Nachweis der
Hormone (Insulin und Glukagon) gelungen (LAXGE 1973, DORN et al. 1975 - dort ausfiihrliche
Literatur). Zur histochemischen Erfassung einer Substanz in del' Zelle odeI' im Gewebe ist ihr Vorhandensein
in e iner bestimmten Konzentration erforderlich. Die Schwierigkeiten der histochemischen Nachweis
verfah ren bestehen darin, die durch Enzymwirkung entstandenen R eaktionsprodukte unloslich zu
machen und kontrastrewh anzufal'ben , um eine lichtmikroskopische Beurte ilung zu ermoglichcn.
Die Problematik del' histochemischen Dat'stellung del' Enzymwirkung ist vielschicht ig.
Nachfolgend werden die Ergebmese der von uns untcrsuchten Enzyme besprochen, zunachst die
Oxidoreduktasen und anschliel3end die Hydrolasen. Die histochemischen Enzymverteilungsmuste ..
im exokrinen und endokrinen Pankreas in Abhangigkeit von del' Ernahrung werden diskutiert und
im Vergleich zur normalen vVistarratte betrachtet. Zu Fragen del' Topochemie del' Enzyme im Pan·
kreas der Sandratte lag uns k ein e Literatur vor. Bei del' Besprechung der r elativen Enzymaktivitiiten werden die jeweiligen Erniihrungsgruppen verglichen.
1. Oxidoreduktasen
1.1. Glukose- 6- phosp hatdeh y dro genase, G6PDH Die G6PDH katalys im·t die Oxidation von d·Glukose-6.phosphat zu 6-Phosphor.£5.gluconolacton.
AlsCoferment fiir diese Reaktion wird NADP verwendet. Die G6PDH ist extramitochondriallokalis iert , \Voraus hervorgcht , dal3 dieses Enzym boi del' konventionellen Tochnik einer nicht unerheb·
lichen Enzymdiffusion ausgesetzt ist. DORX et al. (1970/71) fiihrten in diesem Zusammenhang histo chemi~che Untersuchungen an Kryostatschnitten und vergleichend dazu an isolierten Inseln durch. Mit cineI' Methode von RUDOLPH und KLEIN, modifiziert nach SASSE (P. ARNOLD, 1968) konnten sre in den isolierten Inseln eine hohere G6PDH-Aktivitat naehweisen. Fiir den relativ geringen Aktivi
tatsausfall am Kryostatschnitt wird d ie durch das Kryostatschnittverfahren verletzte Plasmamem
bran vtlrantwortlich gemacht, so daf3 hie I' eine ungehinderte Enzymdiffusion stattfinden kann, wah·
rend die Integl'itat der Zelle bei der isolierten Insel erhalten bleibt.
Be i unseren Untersuehungen konnten wir bei allen Ernahrungsgruppen e ine leichte G6PDH
Aktivitat sowohl in den Inseln als auch im exokrinen Pankreas nach weisen.
Difl G6PDH-Aktivitat in den Langer'hansschen Inseln lag bei del' Basisgruppe und bei den ad
libitum ernahrten Tieren etwas hoher als die des exokrinen Pankreas, so daf3 sich die Inseln yom
exokrinen Pankreas abhoben_ Eine gleichstark e Aktivitat konnten wir lediglich im endo- und exo
krinen Pankreas in den Ernahrungsgruppen < 30 Kcal/d feststellen. Eine J;1oherc Aktivitat del' G6PDH im Inselsystem gegeniiber dem exokrinen Pankreas fanden
GOSSNER (1959), RUDOLPH und KLEIN (1964) und DORN et al. (1970) bei del' Ratte, DORNet al. (1970/ 1971) bei del' Maus und BJORKMANN et al. (1963) beim Pferd. LAZARUS et al. (1964) fiihren die starkere

224 A. DORN und G. KOCH
Aktivitat der G6PDH auf den Glukoseabbau in den Inselzellen uber den Pentosophosphatzyklus zuruck. der glykolytische Abbau soil hier eine untergeordnete Holle spielen. 1m Gegensatz zu den oben genannten Autoren konn ten GEPTS et aI. (1970) am Menschen und LACY (1962) am Kaninchen
histochemisch eine gleichstarke G6PDH·Aktivitat im exokrinen und endokrinen Pankreas nachwei
sen. GEPTS et aI. (1970) untersuchten die G6PDH-Aktivitat auch bei diabetischen Menschen und konnten ein Jeichtes, aber nicht signifikantes Ansteigen der G6PD'H-Aktivitat in den Inseln fest
stellen. Diese Autoren konnten auch registrieren, da/3 eine mutterliche 'Hyperglykamie keinen nennenswerten Einflu/3 auf die Anderung der G6PD'H-Aktivitat in den foetalen Inseln hat.
1.2. Lactatdehydrogenase, LDH
Die Lactatdehydrogenase katalysiert mit NADH2 als Coferment die Hydrierung von Pyruvat zu
Lactat. Die LDH ist extramitochondrial lokalisiert und zeigt besonders starke Aktivitaten in der
Skelettmuskulatur und in der Leber. DE DUVE, W ATTIAUX und BAUDHUIN(1962) geben als intraz eUulare Lokalisation der LDH das Hyaloplasma an, wenngleich eine teilweise Beziehung zum endoplas
matischen Hetikulum von ihnen nicht ausgeschlossen wird. Generell ist die Aktivitat der LD'H im
exokrinen Pankreas starker ausgepragt als in den Langerhansschen Inseln. Untersuchungen dazu
wurden u. a. durchgefiihrt von PETKOV (1969) an verschiedenen Wirbeltieren (Hatte, Meerschwein
chen, Hamster und Katze), DORN et aI. (1970) an der Maus, GEPTS, GREGOIRE, VAN ASSCHE und GASPARO (1970) am Menschen, PETKOV, GALABOWA und GOSPODINOW (1968) am Goldhamster und
DORN et aI. (1973) an der Hatte in prae- und postnatalen Entwicklungsstadien. DORN et aI. (1973)
sahen bei der neugeborenen Hatte eine gleichstarke Aktivitat der LD'H sowohl im exokrinen als auch
im endokrinen Pankreas. Bei unseren Untersuchungen sahen wir in der Basisgruppe und in den Gruppen, die 28 bis 35
Tage mit weniger als 30 Kcal bzw. mehr als 40 Kcal pro Tag ernahrt wurden, einen gleichstarken Ausfall der Enzymreaktion sowohl im exokrinen Pankreas als auch in den Langerhansschen Inseln.
Eine relativ kurzzeitige Ernahrung der Sandratte (14 bis 21 Tage) mit < 30Kcal/d bzw. > 40 KcalJd zeigte ein Verteilungsbild der LDH irn Pankreas, wie wir es von der Wistar-Hatte her kannten, woBei die Heaktion insgesarnt ~tarker war als bei den Wistarratten. In diesen Ernahrungsgruppen fanden wir eine hohere Enzyrnaktivitat in den exokrinen Anteilen des Pankreas. DORN, LIPPMANN und HAHN (1970) fuhrten enzymhistochernische Untersuchungen mit 'Hilfe der konventionellen Technik an isolierten Langerhansschen Inseln durch und fanden eine starke Aktivitat in den Inselzellen. Diese Befunde sind sicherlich auf die erhaltene Integritat der Zellen zuruckzufuhren, die einer Enzyrndiffusion entgegemvirkt.
1.3. ,8-Hydroxibutyratdehydrogenase, HBDH
Die 'HBDH katalysiert die HeaktlOn von Acetonacetat zu p-Hydroxybutyrat, wobei NAD als
Coferment auftritt. Dieses Enzym ist an die Struktur der Mitochondrien gebunden. Die Angaben
zum Nachweis der 'HBD'H waren in der uns zuganglichen Literatul recht sparlich. 'HELLEERSTROM
(1963) konnte im Pankreas der Ente in den Langerhansschen Inseln eine positive EnzymreaktlOn
nachweisen, wahrend sich der exokrine Anteil negativ verhielt. Einen gleichstarken Heaktionsaus
fall in den Langerhansschen Inseln und in den Acmi konnten VAN ASSCHE (1969) und VAN ASSCHE.
GEPTS und DE GASPAO (1969) feststellen. 1m Pankreas von Kaninchen und Hatten kamen GEPTS
und TOUSSAINT (1964) zu negativen Ergebnissen. DORN et aI. (1973) konnten bei neugeborenen Hatten eine zweimal hohere Aktivitat in den A. und B-Zellen nachw81sen als bei adulten Hatten. Die Aktivitat lie/3 jedoch bereits am 2. Tag postnatal nach, und am 5. postnatalen Tag war nur noch eine ma/3ige Aktivitat, wie sie dann auch bei adulten Hatten vorhanden war, zu beobachten. Bei
unseren Untersuchungen an der Sandratte zeigte sieh bei allen Ernahrungsgruppen ein relativ gleichformiges Bild. Die Intensitat des Heaktionsausfalles beim NlWhweis der 'HBD'H war in den Langer
hansschen Inseln und in den Aeini des Pankreas gleich sta,rk a.usgepra€;t. Lediglieh die Ernahrungs
~ruppe. dl'll:" 14 bis 21 Tage > 40 Kcal/Tag gefiittert wurden, z,)igte einl'lI! ()tw~~ 8tarkerell :Reaktions-
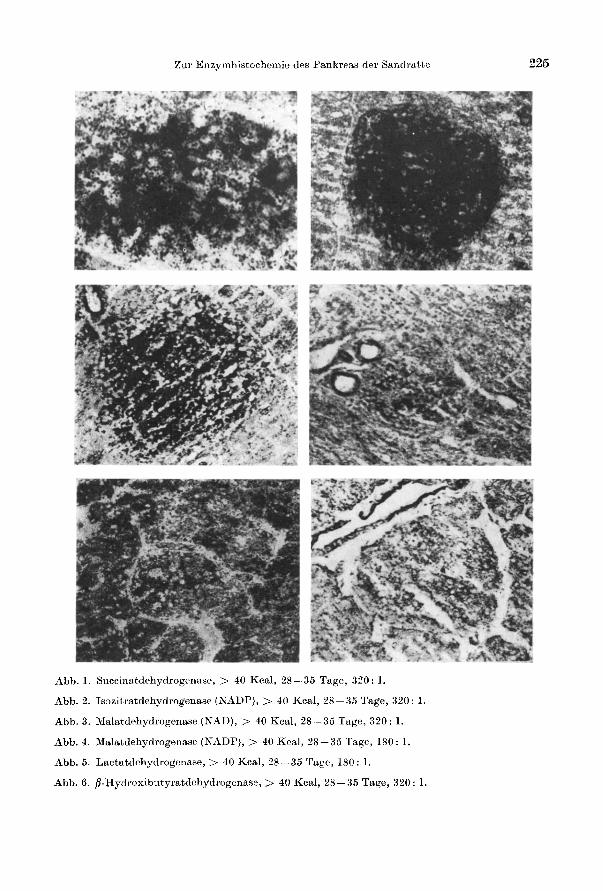
Zur Enzymhistochemie des Pankreas del' Sandratte
Abb. 1. Succinatdehydrogenase, > 40 Kcal, 28 - 35 Tage, 320: 1.
Abb.2. IS:lzitratdehydrogenase (NADP), > 40 Kcal, 28-35 Tage, 320: 1.
Abb.3. }Ialatdehydrogenase (NAD), > 40 Kcal, 28-35 Tage, 320: 1.
Abb.4. Malatdehydrogenase (NADP), > 40 Kcal, 28-35 Tage, 180: 1.
Abb.5. Lactatdehydrogenase, > 40 Kcal, 28-35 Tage, 180: 1.
Abb.6. p·Hydro"ikltyratdehydrogenase, > 40 Kcal, 28-35 Tage, 320: 1.
225
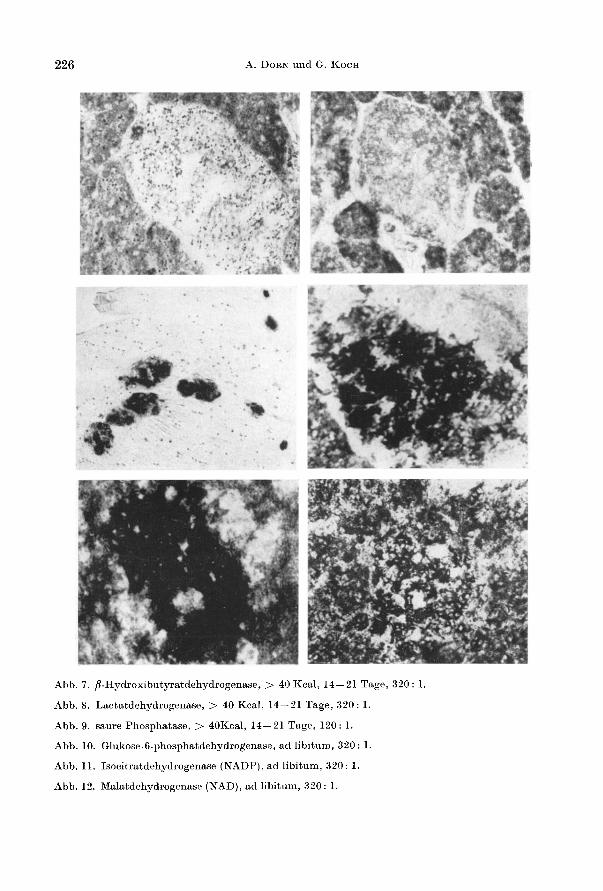
226 A. DORN und G. KOCH
Abb. 7. p.Hydroxibutyratdehydrogenase, > 40 Kcal, 14- 21 Tage, 320 : 1.
Abb.8. Lactatdehydrogenase, > 40 Kcal, 14 - 2l Tage, 320: 1.
Abb. 9. saure Phosphatase, > 40Kcal, l4 - 2l Tage, l 20 : 1.
Abb. lO. GIukose.6-phosphatdehydrogenase, ad libitum, 320: 1.
Abb. li. Isocitratdehydrogenase (NADP). ad libitum. 320: 1.
Abb. 12. Malatdehydrogenase (NAD). ad libitum, 320: 1.
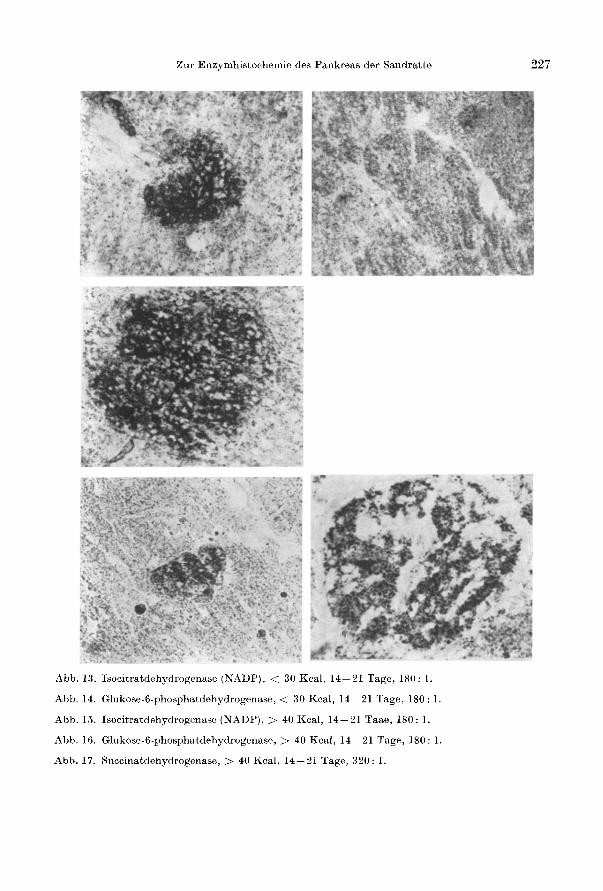
Zur Enzymhistochemie des Pankreas der Sandratte
Abb. 13. Isocitratdehydrogenase (NADP), < 30 Kcal, 14- 21 Tage, 180: 1.
Abb. 14. Glukose-6-phosphatdehydrogenase, < 30 Kcal, 14- 21 Tage, 180: 1.
Abb. 15. Isocitratdehydrogenase (NADP), > 40 Kcal, 14- 21 Taae, 180: 1.
Abb. 16. Glukose-6-phosphatdehydrogenase, > 40 Kcal, 14- 21 Tage, 180: 1.
Abb.17. Succinatdehydrogenase, > 40 Kcal, 14-21 Tage, 320: 1.
227

228 A. DORN und G. KOCH
ausfall im exokrinen Pankrea~. Bei der Emahrungsgruppe, die 14 bis 21 Tage < 30 Kcal/Tag bekam, war allgemein ein schwacherer R eaktionsausfall festzustellen, jed och zeigten die Langerhans
schen Inseln und das exokrine Pankreas gleiche Reaktionsintensita t . Vielleicht kann das Verhalten
dieser Erniihrungsgruppe als eine R eaktion des Organismus auf die vel'iinderte Erniihrungssituation,
die nur relativ kurze Zeit andauerte , angesehen werden. Bei Tieren , die 28 bis 35 Tage auf diese 'Weise erniihrt wurden, konnte sich dieser Effekt wieder zuriickgebildet haben bzw. wurde kompen
siert . Wie auch andere Autoren schon zum Ausdruck brachten, kann man Riickschliisse auf bio
chemische Stoffwechselvorgange durch methodisch bedingte Grenzen der histochemischen Nachweisverfahren nur mit groBer Zuriickhalt ung ziehen. Dieser Einschrankung unterliegen natiirlich
auch unsere Unte rsuchungen .
1.4. Isocitratdehydrogen a se , ICDH (NAD) und ICDH (NADP)
Die NADP-abhangige ICDH bewirkt die Umwandlung des I socitlates iiber Oxalsuccinat zu K etoglutarat. Wir untersuchten die ICDH mit den Cofermenten NAD und NADP. Diese Unter
teilung fanden wir in der uns zur Verfiigung stehenden Literatur allerdings nicht. Einigen Arbeiten
entnahmen wir, daB es sich bei der dort nachgewiesenen lCDH urn ein NADP-abhiingiges Enzym
handelte. Wir hatten Untersuchungen mit beiden Cofermenten durchgefiihrt, da man annimmt, daB die NADP-abhangige lCDH im Grundstoffwechsel der Zelle v erankert ist, wahrend die NAD
abhiingige ICDH einer Stoffwechselregulation der Zelle unterliegen soil. Bei der NADP-abhangigen ICDH fanden wir in der Literatur e ine iibereinstimmende Beurteilung der Enzymaktivitiit. Del'
Reakt ionsausfall war in den Langerhansschen Inseln, hier meist in den B-Zellen, starker als im exo
krinen Pankreasantei1.
WEGMANN und PETKOV (1965, 1966) untersuchten menschlichen Pankreas, VAN ASSCHE, GEPTS und DE GASPARO (1969) und VAN ASSCHE (1969) menschliche Foeten und Neugeborene, PETKOV
et al. (1971) das Schaf, PETKOV (1967) die Katze. GEPTS et a!. (1970) sahen unter Cortisonbehand
lung einen Anstieg der ICDH im Parenchym und in den Inseln be i diabetischen und nichtdiabetischen Patienten. DORN et a1. (1973) konnten bei neugeborenen und adulten Ratten eine hohere Enzymaktivitiit der NADP-abhiingigen ICDH In den Langerhansschen Inseln nachweisen im Vergleich zur schwiicheren Reaktionslage des exokrinen Pankreas. 1m Vergleich zur NAD-abhangigen ICDH erkannte man eine groBere Aktivitiit der NADP-abhangigen ICDH besonders in den A- und B-Zellen. Bei unseren Untersuchungen am Pankreas der Sandratte k onnten wir bei allen Ernabrungsgruppen eine gleich stark ausgepriigte Aktivitat nachweisen. Die Aktivitiiten waren jedoch gegeniiber der Wistarratte besonders in den A-Zellen deutlicher. Die R eaktion fiel in allen Gruppen in den Langerhansschen Inseln starker aus als in den Acini und entsprach somit dem topochemi
schen Verhalten der lCDH im Pankreas der Wistarratte.
1.5. Succinatdehydrogenase, SDH
Die Succinatdehydrogenase katalysiert im Zltratzyklus die reversible Umwandlung von Succinat
in Fumarat. Ein Zusatz von Coenzym im Inkubationsmedium ist nicht erforderlich. Die SDH ist
intramitochondriallokalisiert. Eine zuverlassige Nachweisreaktion der SDH stellt die von PADY
KULA (1952) modifizierte Methode von SELIGMANN und RUTHENBERG dar.
In der Literatur fanden wir fast iibereinstimmend eine hohere Aktivitat im exokrinen Pankreas.
Untersuchungen dieses Fermentes wurden insbesondere durchgefiihrt von : HELLERSTROIII (1963) an
d er Ente, HELLERSTROM und HELLMAN (1962), PETKO V (1968) am Goldhamster, PETKOV (1967) an del' Katze, DoRN et al. (1970) an der Maus, 'WEGMANN et a!. (1965) am Menschen. DORN et a1. (1970/71}
konnten an siolierten Inseln der Maus einen hoheren Reaktionsausfall fes tst ellen im Vergleich zu Ergeb
nissen , die mit Kryostatschnittverfahren erzielt wurden. Der Faktor der Enzymdiffusion durch Ver
letzung der Integritat del' Zelle beim K ryostatschnitt diirfte hierbei wieder eine Rolle spielen. Bei der Ratte fanden CAVALLERO und SOLCIA (1964) in den B-Zellen einen starkeren Reaktionsausfall
als im exokrinen Gewebe. GEPTS und TOUSSAINT (1964) kamen ebenfalls zu diesem Ergebnis. Bei

Zur Enzymhistochemie des Pankl'eas del' Sandratte 22\:1
unseren Untersuchungen am Pankreas del' Sandratte fanden wir in allen Ernahrungsgruppen eine
starke Enzymaktivitat del' Lang81'hansschen Inseln, insbesondere in den B-Zellen, die die Aktivitat des exokrinen Pankresa deutlich iiberstieg. Del' Reaktionsausfall war bis auf die Basisgruppe und
die Tiere, di e 14 bis 21 Tage mit < 30 KcalJd ernahrt wurden, in allen Gruppen von gleicher Intensitat. Die 1e tztgenannte zeigte in A-, B- und exokrinen Pankreaszellen nUl' eine schwache R eak.
tion. Dieses Verha1ten entsprach del' Enzymverteilung beim Hund, wie sie von PETKOV (1967) beobachtet wurde.
1.6. Malatdehydrogenase, MDH (NAD) , MDH (NADP) Die Malatdehydrogenase katalyswrt die oxidative Decarboxylierung von L·Malat und die De·
<larboxylierung von Oxalaceta t . Als Coenzyme verwendeten wir sowohl NAD als auch NADP.
Nach DE DUVE, WATTIAUX und BAUDHUIN (1962) kann die Malatdehydrogenase sowohl in den
Mitochondrien a ls auch im H yaloplasma lokalisiert sein. Die Untersuchungen zur Histotopochemie
del' MDH ergaben bei fast allen untersuchten Tieren eine starke EnzymreaktlOn im endokrinen An·
teil des Pankreas, insbesondere in den B.Zellen, in den Acini war die Reaktion weniger stark ausge· pragt.
WEGMANN und PETKOV (1965), LAZARUS und YOLK (1962) beim Kaninchen, PETKOV et al. (1965)
bei del' Ratte und PETKOV (1967) bei del' Katze kamen ebenfalls zu diesem Ergebnis. Die genannten
Autoren zeigten in ihren Ergebnissen nul' die NAD.abhangige MDH, Hinw<'l ise auf Untersuchungen
del' NADP·abhangigen Enzymaktivitat fehlten in del' uns zuganglichen Literatur.
DORN et a l. (1973) wiesen bei adulten Ratten hohe Aktivitaten in den B·Zellen und im exokrinen Pankreas nach, wahrend die A·Zellen nur einen mal3igen Reaktionsausfall zeigten. Bei neugeboren en Ratten fanden sie in den Acini cine hohere Aktivitat als in den von del' Reaktionsintensitat her
glcichen A· und B- Zellen. In den Entwicklungsstufen bis hin zur Rdulten Ratte konnten sie nul' ein
differentes Verhalten det" MDH nachweiEen. Differenzen in del' nachweisbaren Enzymaktivitat fanden Wlr auch bei den verschledenen Er·
.nahrungsgruppen del' Sandratte . In del' Basisgruppe, die von der Ernahrungslage etwa mit den von
uns untersuchten Wistarratten vergleichbar ist, fanden wir nul' eine schwache MDH.Aktivitat, deren Intensitat in den A ., B - und exokrinen Pa11.kreaszellen gleich war, ebenso verhielt sich das
Tierkollektiv, das 14 bis 21 Tage mit> 40 KcalJd ernahrt wurde. Ein ahnliches Aktivitatsverhalten ze igen ad libitum ernahrte Tle re und die Ernahmngsgruppe > 40 Kcal/d iiber 28 bis 35 Tage. Hie· war ein mal3iger R eaktionsausfall im exokrinen Parenchym und oine deutliche R eaktion del' Langer. hansschen I11.8cln zu verzeichnen, so dal3 sich diese deutlich yom exokrinen Anteil abhoben. Tiere , die mit < 30 KcalJd 14 bis 21 Tage ernahrt wurden, zeigten starke MDH·Aktivitaten im exokrinen Anteil, die Intensitat del' Aktivit,at del' Langerhansschen Inseln war geringer. Wurde dieses Fiitterungsmgime 28 bis 35 Tage belbehalten, EO war ein Reaktionsabfall Bowohl im exokrinen Pankreas ~tls auch in den B-Zellen zu verze ichnen, die Reaktionslage del' A·Zellen blieb unverandert. Ob diese auffalligen Aktivitatsanderungen bei langer dauerndem Diatregime (28 bis 35 Tage) im Zellstoff· wechsel ihre Ursache haben oder ob die Aktivitatsunterschiede methodisch bedingt sind, mul3te
weiteren Untersuchungen vorbehalten bleiben. Die NADP·abhangige Malatdehydrogenase zeigte
in allen Ernahrungsgruppen gleiches Verhalten.
2. Hydrolasen
2.1. Saur e Phosphatase Phosphatason spielen e ine wichtige Rolle beim aktiven Transport durch die Membran. B eson·
del'S die A· und B·Zellen im Pankreas sind als Orte hochster enzymatischer Aktivitiit aufzufasson,
da sie tiber oinen erheblichen Kohlenhydrat. und Proteinum"Xltz ve rfiigen, so dal3 die Untersuchung der sauren Phosphatase uns besonders interessant erschien . Dcr Nachweis del' saUl'en Phosphatase
erfolgte mit e iner Azc~~~ -ustoffmethode. 'Vir verwendeten Echtblausalz BB. HELLERSTROM, BROLIN, LARSSON und HELLMAN (1962) konnten bereits an 20 Tage alten Ratten·
feten cine deutliche s·Pase·Aktivitat des Inselgewebes nanhweisen, wahrend das exokrine Paren·

230 A. DORN und G. KOCH
chym negativ reagierte. In del' postnatalen Entwicklung bls hin zur adulten Ratte konnten dann von diesen Autoren keine Veranderungen im topochemischen Verhalten dieses Enzyms mehr fest
gestellt werden. An der Maus sahen HELLERsTRaM, HELLMAN und TALJEDAL (1964) auch die starke
Enzymreaktiop in den Langerhansschen Inse1n, am Kaninchen wurdo diesel' Befund ebenfalls von
GassNER (1959) und LAZARUS, BARDEN und BRADSHAW (1962) erhoben. PETKOV und DONTcHEv
(1971) sahen am Entenpankreas in den B-Zellen eine sehr starke s-Pase-Aktivitat, wahrend sie in
den A-Zellen und den Acini fehite oder nul' schwach ausfie!' Boi unseren Untersuchungen an del' Sandratte fanden wir sowohl in dol' Basisgruppe als auch
bei Tieren, die 14 bis 21 d mit < 30 Kcal/d ernahrt wurden, eine sehr starke Reaktion in den B
Zellen, in A- und exokrinen Pankreaszellen war keme Enzymaktivitat nachweis bar. Das histotopo
ehemische Bild des Pankraas del' Ernahrungsgruppen 14 bis 21 d > 40 Kcal/d und ad libitum entspraeh
den oben beschriebenen Gruppen. Die Intensitat del' Reaktion in den B-Zellen war jedoch etWas
schwacher. Bei den Gruppen, die die langste Zeit (28 bis 35 d) einem Diatregime unterworfen waren, konnt8n wir keine Aktivitat del' sauren Phosphatase im Pankreas feststellen.
Von einigen Autoren ist das Verhalten der sauren Phosphatase bei diabetischer Stoffwechsellage
untersucht worden. GassNER (1958) gelang der Nachweis del' sauren Phosphatase in den Langerhans
schen Inseln bei jugendlichen Diabetikern nicht. Diesel' Befund wurde als Insuffizienz oder volliger Ausfall del' B-Zellen beim jugendlichen Diabetiker gedeutet.
GEPTS et al. konnten 1970 mit quantitativen Untersuchungen in gewisser Hinsicht die Ergebnisse
G6SSNERS bestatigen, indem sie beim Altersdiabetes im Pankreas noeh eine geringe s-Pase-Aktivitat
fanden. PUTZKE und TESSMANN (1971) konnten am exokrinen Mausepankreas eine Zunahme del'
s-Pase-Aktivitat nach Behandlung mit Testosteron nachweisen.
2.2. Glukose-6-phosphatase, G-6-Pase
Die Glukose-6-phosphatase ist ein spezifisches Enzym del' Glukoneogenese und ist Cofermentunabhangig. Ein Eingrcifen del' Glukose-6-phosphatase in den Mechanismus del' Insulinausschuttung
in den B-Zellen wird diskutiert. TALJEDAL (1969) konnte einen hemmenden EinfluB von Glukose (200 mg/100 ml) auf die G-6-
Pase nachweisen. Da nun hohe Konzentrationen von Glukose hemmend auf die G-6-Pase wirkten, kam es dabei zu einer Anhaufung von Glukose-6-phosphat mit einer nachfolgenden Insulinausschuttung, die als Antwort auf die erhohte Glukosekonzentration angesehen werden kann, so daB die G-6-Pase zu den Schlusselenzymen des Stoffwechsels zahlt und fUr die Funktion del' B-Zellen von be
Bonderer Bedeutung ist. LAZARUS (1959) und G6SSNER (1969) konnten beim Hund eine starke G-6-Pase-Aktivitat in den
Langerhansschen Inseln nachweisen, PETKOV (1966) an del' Katze und PETKOV et a!. (1971) am
Schaf kamen eben falls zu diesem Ergebnis. DORN et al. (1970/71) wiesen die Glukose-6-phosphatase
mit der Methode von WACHSTEIN und MEISEL (1956) an del' Maus nach und sahen in den Langerhansschen Inseln eine Aktivitat dieses Enzym~, wahrend das exokrine Pankreas negativ reagierte. Ein
gleichzeitig durchgefuhrter Nachweis dieses Enzyms an isolierten Inseln brachte eine zweifach hohere Aktivitat. Weiterhin konnten bei diesen Untersuchungen mit langerer Inkubationsdauer ein
Fermentschwund nachgewiesen werden. Bei unseren Untersuchungen an del' Sandratte war ein Nachweis von Glukose-6-phosphatase
Aktivitat in allen Ernahrungsgruppen nicht moglich. Auch bei parallel untersuchtem Pankl'eas del' Wistar-Ratte konnten wir ebenfalls keine Aktivitat ieststellen. In Ubereinstimmung mit unseren
Befunden fanden PETKOV et al. (1968) am Goldhamster, WEGMANN und PETKOV (1965) am Menschen und PETKOV (1969) am Menschen, am Hamster und an del' weiBen Ratte keine G-6-Pase-Aktivitat.
GEPTS et al. (1964) bestatigten ebenfalls das Fehlen einer histochemisch erfaBbaren G-6-Pase. DORN
et al. (1973) konnten in postnatalen Entwicklungsstadien del' Ratte auch keine Aktivitat nachweisen. Urn histotopochemisch G-6-Pase an del' Ratte nachweisen zu konnen, bietet sich die isolierte
Ratteninsel an_

Zur Enzymhistochemie des Pankreas del' Sandratte 231
MAYBAUER (1974) fiihrte vergleichende Untersuchungen zum Nachweis del' Glukose-6-phosphat
ase an Kryostatschnitten von Rattenpankreas und an isolierten Langerhansschen Inseln del' Ratte
dureh_ Die Reaktion war am Sehnitt durch das Gesamtpankreas negativ. An del' isolierten Langer
hansschen Insel del' Ratte konnte er mit Hilfe einer Ca-Methode eine gut naehweisbare Aktivitat del' G-6-Pase zeigen.
2.3. Adenosintriphosphatase, ATPase
Die ATPase spaltet die letzte Anhydridbindung des ATP. Es wird die Moglichkeit diskutiert,
dal3 eine del' Funktionen del' ATPas3 darin besteht, eine ATP-Anhaufung zu verhindern und konti
nuierlieh ATP fUr den Fortgang del' Stoffweehselvorgange zur Verfiigung zu stellen. Die ATPase
wurde in Mikrosomen, in Mitochondrien und an del' Zellmembran nachgewiesen.
Man mul3 u. a. eine Mg++- und (oder) Ca++-aktivierte ATPase unterscheiden. Ihr Wirkungs
optimum ist abhangig YOm pH-Bereich. Die ATPase-Aktivitat wurde oft histotopoehemisch unter
sucht, und zu ihrem Nachweis wurden in den letzten Jahren mehrere Verfahren entwickelt, die die
ATPasen bei unterschiedlichen pH-Werten und verschiedenen Aktivatoren darstellen. Zum Nachweis
del' ATPase benutzten die meisten Autoren entweder die Bleisalzmethode nach W ACHSTEIN und
MEISEL (1957) (pH = 7,2) odeI' die succedane Calcium-Methode nach PADYKULA und HERMAN.
Histochemische Untersuchungen mit del' Methode nach W ACHSTEIN und MEISEL ergaben bei DORN
et el. (1970/71) an del' Maus und bei PETKOV und DONTCHEV (1971) am Entenpankreas eine Lokalisa
tion del' n-ATPase in den Langerhansschen Inseln, insbesondere in den B-Zellen. An isolierten
Inseln und am Paraffinschnitt zeigte die n-ATPase eine intensivere Reaktion als in den Kryostat
schnitten. DORN et al. (1974) sahen bei Wistar-Ratten in allen von ihnen untersuchtcn postnatalcn
Entwicklungsstufen keine n-ATPa~e-Aktivitat, die sie auf eine eventuell besonders intensive Enzym
diffusion del' n-ATPase im Kryostatschnitt zuriickfiihrten.
Bei unseren Untersuchungen war in allen Ernahrungsgruppen eine mal3ige Aktivitat del' n-ATP
ase sowohl im exokrinen Parenchym als auch in den Langerhansschen Inseln nachweisbar. Ernah
rungsgruppen, die 28 bis 35 Tage einem Diatregime von < 30 Kcal/d bzw. > 40 Kcal/d unterworfen
waren, zeigten in den Inseln sogar eine gut nachweisbare Enzymaktivitat gegeniiber einer mal3igen
Reaktion im exokrinen Pankreasanteil. Aus del' Reaktionsweise der einzelnen Ernahrungsgruppen
geht hervor, daB fiir einen negativen Ausfall der Nachweisreaktion del' ATPase nicht allein die Enzymdiffusion bei Kryostatschnittechnik verantwortlich gemacht werden kann, sondern noch andere Ursachen vorliegen miissen.
Ein Nachweis del' alkalischen ATPase erfolgte von PETTERSON (1966), WEGMANN et al. (1967)
am Meerschweinchen, WEGMANN und PETKOV (1966), PETKOV (1971) am Schaf, DORN et al. (1970,
1971) an del' Maus und DORN et al. (1976) an neugeborenen und postnatalen Entwicklungsstufen
del' Ratte. Besonders am isolierten Inselpraparat und am Paraffinschnitt konnten DORN et al. (1971) Kapillarwande mit einer schr hohen ATPase-Aktivitat nachweisen.
Uns ist del' histochemische Nachweis del' alkalischen ATPase in keiner Ernahrungsgruppe del'
Sandratte gelungen, was abel' nicht zu del' Schlul3folgerung fiihren sollte, dal3 dieses Enzym nicht
vorhanden ist. Um eindeutige Aussagen treffen zu konnen, Ware die Untcrsuchung grol3erer Tier
kollektive notwendig.
Literatur
ARNOLD, M., Histochemie. Einfiihrung in Grundlagen und Prinzipicn del' Methoden. Springer Verlag
Berlin-Heidelberg-New York 1968.
BARMAN, T. E., Enzyme handbook Vol 1 und 2. Springer Verlag, Berlin, Heidelberg, New York 1969.
BJORKMAN, N., HELLERSTROM, C., HELLMAN, B., and ROTHMANN, U., Ultrastructure and enzyme
histochemistry of the pancreatic islets in the horse. Z. Zellforsch. 59, 535-554 (1963).
CAVALLERO, C., and SOLCIA, E., Cytologic and cytochemical studies on the pancreatic islets. In:
The structure and metabolism of the pancreatic islets. p. 83 - 97, Pergamon Press Oxford 1964.

232 A. DOR:S und G. KOCH
DE DUVE, C., WATTIAUX, R. and BAUDHl:IN, P., Distribution of enzymes between subcellular fractions in animal tissues. Advanc. Enzymo!. 24, 291-358 (1962).
DORN, A., lIAHN, 1I .. J., LIPPMANN, 1I .. L., MICHAEL, R., und LIPPMANN, H. G., Untersuchungen
an Langerhansschen Inseln in vitro. IV. Mitteilung. Acta histochem. 43, 41- 50 (1972).
KOCH, G., NISSEN, L., SCHARKOWSKI, K. R., GRUSS, G., MAYBAUER, W., und FEHRMANN, P.,
Enzymhistochemische Untersuchungen an Langerhansschen Inseln der Wistar·Ratte wahrend
der postnatalen Entwicklung. I. Mitteilung: Nachweis von Oxidoreductasen. Acta histochem.
46,79-100 (1973).
- HAHN, 1I. J., ZIEGLER, M., Elektronenoptische Untersuchungen zum Verhalten isolierter
Langerhansscher Inseln in vitro nach Behandlung mit reinen isolierten Insulinantikorpern. Endo·
krinologie 61/1,144-148 (1973).
LIPPMANN, 'fL-L., HAHN, 1I. J., Untersuehungen an Langerhansschen Inseln in vitro. II. Mit
teilung. Acta histochemica 36, 368-375 (1970).
SCHAROWSKI, K. R., NISSEN, L., LORENZ, D., KOCH, G., und ZIEGLER, M., Immunhistochemischer
Nachweis von Insulin, Glukagon und Gastrin in den Langerhansschen Inseln del' Wistar-Ratte
wahrend del' Entwicklung. Acta histochem. 56, 270-284 (1976).
FERNER, 1I., Das Inselsystem des Pankreas. Georg Thieme Verlag Stuttgart 1952.
FRENKEL, G., KRAICER, P. F. and SHANI, J., Diabetes in the Sand-Rat: Diabetogenesis, Responses
to Mannoheptulose and Atriplex Ash. Diabetologia 8, 313-318 (1972).
GEPTS, W., GREGOIRE, F., VAN ASSCHE, A., and DE GASPARO, M., Quantitative Enzyme Pattern
and Insulin Content of lIuman Islets of Langerhans. in: The Structure and Metabolism of the
Pancreatic Islets, 283 - 303, Pergamon Press, Oxford 1970.
and TOUSSAINT, D., Effect of cortisone, growth hormone and hypophysectomy on the enzymatic
activity of the pancreatic islets. in: The Structure and Metabolism of the Pancreatic Islets, 357
- 377, Pergamon Press (1964).
GEVER, G., Ultrahistochemie, 2. Auf!. VEB Gustav Fischer Verlag, Jena 1973.
GOSSNER, W., lIistochemischer Nachweis hydrolytischer Enzyme mit lIilfe der Azofarbstoffmethode.
Untersuchungen zur Methodik und vergleichenden lIistotopik del' Esterasen und Phosphatasen
bei Wirbeltieren. lIistochemie 1, 48-96 (1959).
Die vergleichende Morphologie del' Langerhansschen Inseln. lIistochemie del' Langcrhansschen
Inseln. in: PFEIFFER, E. F.: lIandbuch des Diabetes mellitus Pathophysiologie und Klinik. Bd. 1,
63-87 (1969). J. F. Lehmanns Verlag, Miinchen.
HACKEL, D. B., FROHMANN, L., MIKAT, E., LELOVITZ, H. E., SCHMIDT-NIELSEN, K. and KINNEY,
T. D., Effect of diet on the glucose tolerance and plasma insulin levels of the sand-rat. Diabetes
15,105-111 (1966).
SCH)UDT-XIELSEN, K., HAINES, K., and MIKAT, E., Diabetes mellitus in the Sand-Rat. Puthologi.
cal studies. Lab.Invest, 14, 200-207 (1965).
lIAHx, 1I. J., WAPPLER, E., und FIEDLER, R., Die Sandratte (Psammomys obesus): Ein an Spontan
diabetes erkrankendes Tier. Z. Versuchstierkd. 13,275-282 (1971).
HAHN YOX DORSCHE, 1I., FEHRMANN, P., SULZMANN, R., und KRA1:SE, R., Morphologische Ver
anderungen am Sandrattenpankreas vor Diabetesmanifestation. In: "VIII Karlsburger Sympo
sium uber Diabetesfragen, 5. bis 8. Juni 1974". lIerausgeber: H. BIBERGEIL, 1I. FIEDLER und
U. POSER, Karlsburg 1975.
HELLERSTROM, C., Enzyme histochemistry of the pancreatic islets in the duck with special reference
to the two types of A-cells. Z. Zellforschung. 60, 688-710 (1963).
BROLIN, S., LARSSON, S., and lIELLMAX, B., Acid phosphatase activity in the pancreatic islets
of rats of different agens. Aeta Endocrin. 40, 604-612 (1962).
and HELLMAN, B., Demonstration of oxidative enzymes in the human pancreas with special
mference to the islets of Langerhans. Acta path. microbio!' scand. 55, 385-394 (1962).

Zm Enzymhistochemie des Pankreas der Sandratte 233
- PETERSON, B., and HELLMAN, B., Some properties of the B·cells in the islets of Langel'hans Rtudies with regard to the position of the cells. Acta Endocrin. 34,449 - 456 (1960).
KARLSON, Kurzes Lehrbuch del' Biochemie. S. Auflage , Georg Thieme Verlag Stuttgart 1972. KOHLER, E ., HAHN, H. J ., K NOSPE, S., SCHAFER. H., und ZIEGLER, B. , Dic Sandrattc - ein Tip,. ·
modell zur Charakterisierung pradiabetischer Stoffwechselverandel'ungen. In: "VIII. Karlsburger Symposion uber Diabetesfragen 5. bis 8. Juni 1974", S. 58-62.Herausgebel': H. BIBERGEIL, H . FIEDLER und U. POSER. Karlsburg 1975 S. 5S-62.
LACY, P. E. , Quantitative histochemistry of the islets of Langerhans . Diabetes n , 96-100 (1962) .
I.A~IGE, R. R ., Zm Histochemie und Elektronenmikroskopie del' Langerhansschen Inseln. Acta histochem. Suppl. n , 69 - 71 (1971).
- Histochemistry of the islets of Langerhans. In Hdb. del' H istochemie. Hrsgg. von W. GRAUM A:>N
und K. NEUMANN. VIII/l suppl. G. Fischer Verlag Stuttgart 1973.
LAZARUS, S. S., BARDEN, H., and BRADSHAW, :vr., Pancreatic beta cells and alloxan toxicity. Enzymatic histochemistry and toxic mechanism. Arch. Path. 73,210- 222 (1962).
- and VOLK, B. W., The pancreas in human and experimental diabetes. Gruie and Stratton , New York and London 1962.
LIKE, A. A., and MIKI, E., D iabetic syndrom in sand rats. IV. Morphologic changes in islet tIssue. Diabetologia 3, 143-166 (1967).
MAYBAUER, W., Untersuchungen zur histochemischen VerteiLung hydrolytisch er und proteolyti
scher F ermente in del' isolierten Langerhansschen Insel del' we ilJen Wistar-Ratte und ihre Bedeutung im Insulinstoffwechsel. Med. Diss., Greifswald 1974.
MIKI, W., LIKE, A. A., STEINKE, J., and SOELCHXER, J. S., Diabetic syndrome in sand rats II. Varia
bilityand assoziation with diet. Diabetologia 3,135-139 (1967). PEARSE, A. G. E., Histochemistry. Theoretical and Applied. Vol. 2. Churchill Livingstone, Edin
burgh and London 1972.
PETKOV, P. E ., Histochimie du pancreas du chat avec discussion du tissu Langerhans endocl'ine. Histochemie II, 305 -31S (1967).
Histochemie del' Lang'erhansschen Inseln des Pankreas bei den Saugetieren. Anat. Anz. 124,
741 - 750 (1969).
GALABOWA, R., and GOSPODINOV, C., Histochemistry of the pancreatic islets in golden hamster, Mesocrice tus auratus Waterhouse. Histochemie 15, 31S-332 (1968). GOSPODINOV, C., GALABOWA , R., and OGNEVA, V., Recherches histologiques et histochimiqucs des ilots de Langerhans du pancreas du porco Acta diabet. lat. 8, 311-330 (1971).
PETTE, D., und BRANDAU, R., Enzym-Histiogramme und Enzymaktivitatsmuste r del' Rattenleber. Nachweis pyridinnukleotidspezifischer Dehydrogenasen in Gelschicht-Verfahren. Enzym. bioI. clin. 6, 79-122 (1966).
PETTERSON, B. , Cytochemica l studies of h ydrolytic and oxidative enzymes in sections and isoLated cells, from the endocrine pancreas of guinea pigs. Histochemie 7 , 116 - 131 (1966).
PRANGE, H. D., SCHMIDT-NIELSEN, K. N., and HACKEL, D. B., Care and beeding of the fat sand rat.
Lab. Animal. Care 18,170-179 (196S).
PUTZKE, H. P., und TESSMANN, D., Histochemisches Verhalten intermediarer und exportabler En
zyme des endokrinen Mausepankreas unter Testosteronwirkung. Acta histochem. 41 , 7 -12 (1971).
RUDOLPH, G., Histochemie del' Glukose-6-phosphat-dehydrogenase im R ahmen des Pentose-phos
phat-Zyklus. Acta histochem. Suppl. IV, 70 - S1 (1964).
STOWARD, P . J., Fixation in Histochemistry. Chapman and Hall, London 1973.
'l'ALJEDAL, I. B., Presence, induction and possible role of glucose- 6-phosphatase in mammalian
pancreatic islets. Biochem. J. 114,387- 394 (1969). Direct fiuorophotometric recording of enzyme kine tics in cryostat sections. Glucose-6-phosphate dehydrogenase in the endocrine mouse pancreas. Histochemie 21, 307 -313 (1970).
VAN ASSCHE, W., Histoenzymatical study of the fetal pancreas in late pregnancy. BioI. Neonat. 14,
19-27 (1969).
16 Acta hi stochem . Bd. 57

234 A. DORN und G. KOCH
VAN ASCHE, W., GEPTS, W., and DE GASPARO, M., The endocrine pancreas in anencephalus. A histological, histochemical and biological study. BioI. Neonat. 14,374-388 (1969).
WACHSTEIN, M., and MEISEL, E., On the histochemical demonstration of glucose·6.phosphate. J.
Histochem. Cytochem. 4,592-600 (1956). WEGMANN, R., and PETKOV, P., Histoenzymologie du pancreas endocrine de l'homme. Ann. Histo
chim. 10,93-100(1965). WENK, H., RITTER, J., und MEYER, U., Beitrag zum histochemischen Nachweis Pyridin·nucleotid
abhangiger Dehydrogenasen: eine halbquantitative Untersuchung. Acta histochem. 38, 278-
292 (1970).
Anscbrift del' Verfasser: p. A. Doz. Dr. sc. med. A. DORN, Anatomisches Institut del' Universitat
Greifswald, DDR - 22 Greifswald, Fr.-Loeffler-StraJ3e 23c.





![3 Pankreas Löser [Kompatibilitätsmodus] · Darm - Ernährung kritisch Kranker Souba et al. ( 1988 ), Baskin et al. ( 1992 ), Sigurdsson et al. ( 1997 ) größte Grenzfläche zur](https://img.pdfslide.org/doc/110x75/5e1b026682064b30a13b86d5/3-pankreas-lser-kompatibilittsmodus-darm-ernhrung-kritisch-kranker-souba.jpg)






















