Embed Size (px)
Citation preview

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
Verunreinigungen zurückzuführen sind, sondern den Besitz von gleichen oder ähnlichen Antigenen an-zeigen. Keine Aussage ist möglich über die Antigene der Lysosomen, da diese Partikel, wegen der Schwie-rigkeit, sie frei von Mikrosomen-Material zu erhal-ten, nicht in den Antigenvergleich einbezogen wur-den.
Der Nachweis von fraktionsspezifischen, nur in bestimmten Zellorganellen auftretenden Antigenen steht in Übereinstimmung mit den Ergebnissen von P E R L M A N N und D ' A M E L I O an Desoxycholat- und Athylendiaminotetraessigsäure-Extrakten aus Cyto-plasmapartikeln 8.
Gewebespezifische Mikrosomen-Antigene fehlten
4 P. K. VOGT, Z . Naturforschg., im Drude. 5 G. H . HOGEBOOM, W. SCHNEIDER U. M. J . STRIEBICH, J . biol.
Chemistry 196, 111 [1952]. 6 C . DE D U V E , C . PRESSMAN , R . GIANETTO , R . WATTIAUX U . F .
APPELMANS , Biochem. J . 6 0 , 604 [1955]; E. L. KUFF, G . H .
HOGEBOOM U. A . J . DALTON, J . Biophys. Biochem. Cytol. 2 ,
33 [1956],
in Kernen, Mitochondrien und löslichem Cytoplasma-extrakt. Sie gehören zu den fraktionsspezifischen Antigenen. Gewebespezifität und Fraktionsspezifität gehen bei den Lebermikrosomen auf die gleichen Antigene zurück. Es bleibt jedoch die Möglichkeit offen, daß andere Zellfraktionen eigene gewebe-spezifische Antigene besitzen. Die Existenz von be-sonderen Mitochondrien-Antigenen geht aus Tab. 1 hervor. In diesem Zusammenhang ist die Entdek-kung eines Phosphatids von Interesse, das nur in den Mitochondrien gefunden wird9. Eigene, noch unvollständige Versuche zeigen, daß sich die Mito-chondrien der Leber von denen der Niere immuno-logisch unterscheiden lassen.
7 F. APPELMANS , R. W A T T I A U X U. C. DE D U V E , Biochem. J . 5 9 ,
438 [1955]. 8 P . PERLMANN U. V. D 'AMELIO , Nature [London] 1 8 1 , 491
[1958]. 9 G. V. MARINETTI, J . ERBLAND U. E . STOTZ, J . biol. Chemistry
233, 562 [1958],
Die Immunologie der Lebermikrosomen III. Die Lokalisation gewebespezifischer Antigene innerhalb der Strukturkomponenten
des endoplasmatischen Reticulums
V o n P E T E R K . V O G T *
Aus dem Max-Planck-Institut für Virusforschung, Tübingen ( Z . Natur forschg . 15 b, 2 2 1 — 2 2 5 [1960] ; e i n g e g a n g e n am 9. Januar 1960)
Microsomes were fractionated by suspending them in a medium composed of 0.25 M sucrose, 0.08 M sodium citrate, and 0.01 M phosphate buffer pH 7.2, followed by a differential centrifugation. Three fractions were obtained. The first, sedimenting at an Smin of 200 consisted predominantly of the membranous component of the endoplasmic reticulum. Lipoprotein was enriched in this fraction. A second fraction ranging between 20 and 200 Svedberg units contained 70 per cent of the microsomal RNA and included mainly the ribosomes. However, membranous material was found still to be present. The remaining supernatant formed the soluble microsomal extract. Extraction of the microsomes with a fluorocarbon yielded pure ribosome preparations consisting of 60 per cent RNA and 40 per cent pro-tein. These ribosomes existed in two sizes, sedimenting at 70 and 100 Svedberg units. Among the microsomal subtractions prepared by the use of citrate the membranous material showed the highest concentration of tissue specific antigens, followed by the unfractionated microsomes and the ribo-nucleoprotein fraction. No tissue specific antigens were found in the soluble microsomal extract. The residual activity of the ribonucleoprotein fraction was due to contaminating microsomal membranes; pure ribosomes prepared by the fluorocarbon method failed to give precipitates with tissue specific antibody. It is concluded that the tissue specific antigens of the microsome fraction are occuring exclusively in the membranes of the endoplasmic reticulum.
P O R T E R und P A L A D E haben mit elektronenmikro-skopischen Aufnahmen von ganzen, äußerst flach ausgebreitet wachsenden Gewebekulturzellen und von Dünnschnitten die weite Verbreitung eines netz-
* Mit Unterstützung der S t u d i e n s t i f t u n g d e s d e u t s c h e n V o l k e s .
1 K . R. PORTER, J . exp. Medicine 97, 727 [1953] ; Harvey Lect. 5 1 , 175 [1957]; G. E. PALADE U. K . R . P O R T E R , J .
artigen Systems zusammenhängender Gefäße im Cytoplasma verschiedener Zelltypen gezeigt und da-für die Bezeichnung „endoplasmatisches Reticulum" vorgeschlagen 1. In diesem Begriff sind verschiedene
exp. Medicine 1 0 0 , 641 [1954]; G. E. PALADE, J . Biophys. Biochem. Cytol. 2, Suppl. 85 [1956]; 1, 567 [1955],

Organelle wie Ergastoplasma und G o 1 g i - Apparat zusammengefaßt, da sie ein einheitliches Membran-system bilden, durch welches das Cytoplasma ge-kammert wird. Die Aufgabe dieser Kammerung ist nicht bekannt. Dagegen wurden in den Membranen selbst einige Enzyme lokalisiert, wie Glucose-6-Phos-phatase, Cytochrom-c-Reduktasen und sehr wahr-scheinlich auch das mikrosomenspezifische Cyto-chrom m2 . Die den Membranen stellenweise auf-sitzenden Ribonucleoproteid-Partikel oder Riboso-men sind der hauptsächlichste Ort cytoplasmatischer Proteinsynthese3-7. Aus Rattenleber-Homogenaten läßt sich eine Mikrosomenfraktion gewinnen, die zum größten Teil aus Fragmenten des endoplasma-tischen Reticulums besteht 4. Diese Mikrosomenfrak-tion enthält gewebespezifische Antigene, deren Loka-lisation innerhalb der Strukturkomponenten des endoplasmatischen Reticulums in den folgenden Ex-perimenten untersucht wird.
Material und Methoden
1. Immunologie. Die Gewinnung und Absorption von Antiseren und die Durchführung des Präzipitintests sind bereits beschrieben worden 5.
2. Die Präparation der Mikrosomen. Sie wurde nach der ebenfalls bereits in 1. c.5 angegebenen Methode durchgeführt.
3. Chemische Analysen. Das Verfahren von L O W R Y
und Mitarbb. 6 diente zur Proteinbestimmung. Die Ri-bosenucleinsäure wurde mit 0,5-m. Perchlorsäure aus den Mikrosomen extrahiert und dann photometrisch ge-messen 7. Die Lipidextraktion wurde nach den Angaben von S C H N E I D E R 8 durchgeführt, und der Phosphor in den Extrakten nach E R N S T E R und Mitarbb. 9 bestimmt.
4. Elektronenmikroskopie. Mikrosomen und Mikro-somen-Subfraktionen wurden als Ausstrichpräparate in einem AEG Zeiss Elektronenmikroskop geprüft. Sie waren in Osmiumdampf fixiert, in manchen Fällen mit Platin Rhodium unter einem Winkel von 25 Grad be-dampft.
Ergebnisse
1. F r a k t i o n i e r u n g der M i k r o s o m e n . Um die beiden morphologischen Hauptkomponenten
2 T. HULTIN , Exp. Cell Res. 12, 290 [1957] ; P. STRITTMATTER
u. S. F . VELICK , J. biol. Chemistry 221, 253 [1956], 3 P . C . ZAMECNIK, E . B . KELLER, J . W . LITTLEFIELD, M . B .
HOAGLAND U. R . B . LOFTFIELD. J . cellular comparat. Physiol. 47, Suppl. 1. 81 [1956].
5 P. K. VOGT, Z . Naturforschg. 15b. 2 1 3 [ I 9 6 0 ] , 6 O . H . LOWRY , N . J . ROSEBROUGH, A . L . FARR U. R . J . R A N -
DALL, J . biol. Chemistry 193, 265 [1951]. 7 J . W . LITTLEFIELD, E . B . KELLER, J . GROSS U. P . C . ZAMECNIK,
J. biol. Chemistry 217, 111 [1955],
des endoplasmatischen Reticulums, Lipoproteid-Membranen und Ribosomen, voneinander zu tren-nen. können zwei Wege beschritten werden. Die Membranen lassen sich durch Netzmittel auflösen und die Ribosomen aus einer solchen Präparation abzentrifugieren 7 '10 . Oder es werden die Ribosomen durch Ca- und Mg-Entzug von den Membranen ab-gelöst und beide Komponenten ebenfalls durch frak-tioniertes Zentrifugieren getrennt4'11. Der Gebrauch von Netzmitteln ist für immunologische Zwecke nicht geeignet. Desoxycholat-Extrakte von Mikrosomen sind zwar mit Antiserum präzipitierbar; wird jedoch die Antikörperbindungs-Kapazität soldier Präparate gemessen, so ist diese besonders für gewebespezifi-sche Antikörper stark herabgesetzt. Das zeigt, daß ein großer Teil der antigenen Gruppen durch das anhaftende Desoxycholat blockiert wird. Durch Dia-lyse läßt sich die ursprüngliche Antikörperbindungs-Fähigkeit nur teilweise wiederherstellen.
Für die Trennung von Ribosomen und Membra-nen durch Ca-Bindung erwies sidi Citrat als beson-ders gut geeignet. Die Mikrosomen wurden in einer Konzentration von 0,5 mg N-Protein/ml in einem Medium suspendiert, das aus 0,25-m. Saccharose, 0,08-m. Natriumeitrat und 0,01-m. S ö r e n s e n -Phosphatpuffer pn 7,2 bestand. Nach 20 Min. bei 0 °C folgte eine Zentrifugation ps = 25 • 108, 30 Min., kleinste sedimentierte Partikel = 200 Svedberg Ein-heiten. Hierbei wurden die Mikrosomen-Membranen
Protein Ribosen-nucleinsäure
I 1 1 1 I I
Mikrosomen-Membran-fraktion Ribonucleoproteid-Fraktion Löslicher Mikrosomen-Extrakt
82,0
5,8
12,2
52,0
20,6
27,8
82,5
13,2
4,3
10,6
72,3
17,1
Tab. 1. Prozentuale Verteilung von Ribosenucleinsäure und Protein in Mikrosomensubfraktionen (die nicht fraktionierten Mikrosomen = 100%). I = in Saccharose-Medium ohne Citrat.
11 = Saccharose-Medium mit 0,08-m. Citrat.
8 W. C. SCHNEIDER, J . biol. Chemistry 161, 293 [1945]. 9 L . ERNSTER, R. ZETTERSTRÖM U. O . LINDBERG , Acta chem.
scand. 4, 942 [1950], 4 G. E. PALADE U. P . SIEKEVITZ, J . Biophys. Biochem. Cytol.
2, 171 [1956], 10 P. COHN U. J . A. V. BUTLER , Biochim. biophysica Acta [Am-
sterdam] 25, 222 [1957]. 1 1 K . K . TSUBOI , M. DE TERRA U. P. B. HEIDSON , Exp. Cell. Res.
7, 32 [1954]; H . S A C H S , J . biol. Chemistry 233.643 [1958],

Tab. 2. Konzentration von Ribosenucleinsäure, Protein und Phospholipid in Mikrosomen-Subfraktionen. I = Saccharose-Medium ohne Citrat. II = Saccharose-Medium mit 0,08-m. Citrat. * In Prozent von Ribosenucleinsäure plus Protein.
** In ug Phospholipid-P pro mg Protein.
Ribosenucleinsäure * Protein * Phospholipid ** I II I II I II
Mikrosomen Membranfraktion 9,6 2,3 90,4 97,7 66 105
Ribonucleoproteidfraktion 10,0 38,0 90,0 62,0 62 50 Löslicher
Mikrosomenextrakt 6,2 14,0 93,8 86,0 19 25
ausgeschleudert. Aus dem Uberstand läßt sich ein Material abzentrifugieren, das vorwiegend aus Ribo-somen besteht: p; = 66' 108, 120 Min., kleinste sedi-mentierte Partikel = 20 S. Es entstehen so aus den Mikrosomen drei Fraktionen: eine Membranfrak-tion, eine Ribonucleoproteid-Fraktion und ein lös-licher Mikrosomenextrakt. Die Werte der chemischen Analysen für ein typisches Experiment sind in Tab. 1 und 2 wiedergegeben und Kontrollen gegenüber-gestellt, bei denen das Citrat im Medium weggelas-sen wurde.
Durch die Suspension im Citratmedium bleiben 50% des Proteins und 90% der Ribosenucleinsäure bei der ersten Sedimentation im Uberstand. Die ent-sprechenden Werte für die Kontrolle sind 18% für Protein und Ribosenucleinsäure (Tab. 1). Die Mem-branfraktion enthält nur wenig Ribosenucleinsäure, die mit 38% einen Hauptbestandteil der Ribonucleo-proteid-Fraktion bildet. Phospholipide sind in der Membranfraktion angereichert, doch enthält auch die Ribonucleoproteid-Fraktion signifikante Men-gen.
Im Elektronenmikroskop lassen sich in der Mem-branfraktion im Gegensatz zu den intakten Mikro-somen nur wenige Ribosomen erkennen. In der Ribonucleoproteid-Fraktion finden sich neben Ribo-somen auch Membranfragmente, deren Anwesenheit auch durch den Phospholipidgehalt dieser Fraktion angezeigt wird.
Zur Darstellung von Ribosomen, die völlig frei von der Membrankomponente sind, diente eine Methode, die für die Reinigung von RNS-Viren ent-wickelt wurde12. Zu Mikrosomen, suspendiert in einem Medium, bestehend aus 0,25-m. Saccharose, 0,025-m. KCl, 0,005-m. MgCl2 und 0,05-ro. Tris-PufTer pn 7,6, wurde ein halbes Volumen Trichlor-trifluoräthan * gegeben und die Mischung in einem Starmix bei 35 000 U/min unter Kühlung 1 Min. lang homogenisiert. Wäßrige und organische Phase konnten anschließend durch Zentrifugation 10 Min.
3500 U/min getrennt werden. Die wäßrige Phase wurde abpipetiert, erneut Saccharosemedium (ohne Mikrosomen) zur organischen Phase zugegeben und die Homogenisation und Phasentrennung wiederholt. Die gepoolten wäßrigen Phasen wurden nochmals mit einem halben Volumen Trichlortrifluoräthan homo-genisiert, anschließend von der organischen Phase abgetrennt und bei einem p; = 66 -10 8 , 120 Min., Smjn = 20, ausgeschleudert. Das Sediment bestand zu 60% aus Ribosenucleinsäure und zu 40% aus Protein. In Ausstrichpräparaten waren mit dem Elektronenmikroskop gleichförmige Partikel von etwa 20 m/< Durchmesser zu sehen. Die Ultrazentri-fugen-Analyse ergab zwei scharfe Gradienten mit 520 Werten von 70 und 100. Das Verhältnis der UV-Absorptionen 260 m[i zu 280 mja war 1,6.
2. Die V e r t e i l u n g g e w e b e s p e z i f i -scher A n t i g e n e in den M i k r o s o m e n -S u b f r a k t i o n e n .
a) Versuche mit direkter Präzipitation. Werden zwei Stoffgemische, die verschiedene Konzentratio-nen des gleichen Antigens enthalten, mit der quanti-tativen Präzipitinmethode (konstante Menge Anti-serum, ansteigende Mengen Antigen) verglichen, so erreicht die Präparation mit dem relativ höheren Antigengehalt den Äquivalenzpunkt eher. Die Be-stimmung des Äquivalenzpunktes gibt daher Auf-schluß über die Konzentration von Antigenen in einem Material, das daneben noch nicht reagierende Proteine enthält. In Abb. 1 ist eine vergleichende Äquivalenzpunkt-Bestimmung von Mikrosomenmem-bran-Fraktion und nicht fraktionierten Mikrosomen mit gewebespezifischem Mikrosomen-Antiserum wie-dergegeben. Von der Membranfraktion wird weni-ger Protein für die Erschöpfung des Serums benö-tigt als von den Mikrosomen. Die Konzentration der gewebespezifischen Antigene, bezogen auf Protein,
12 A. E . GESSLER, C H . E . BENDER U. M. C . PARKINSON , Trans. N. Y. Acad. Sei. Ser. II, 18, 701 [1956],
* „Arcton 63" Imperial Chemical Industries Ltd., London.
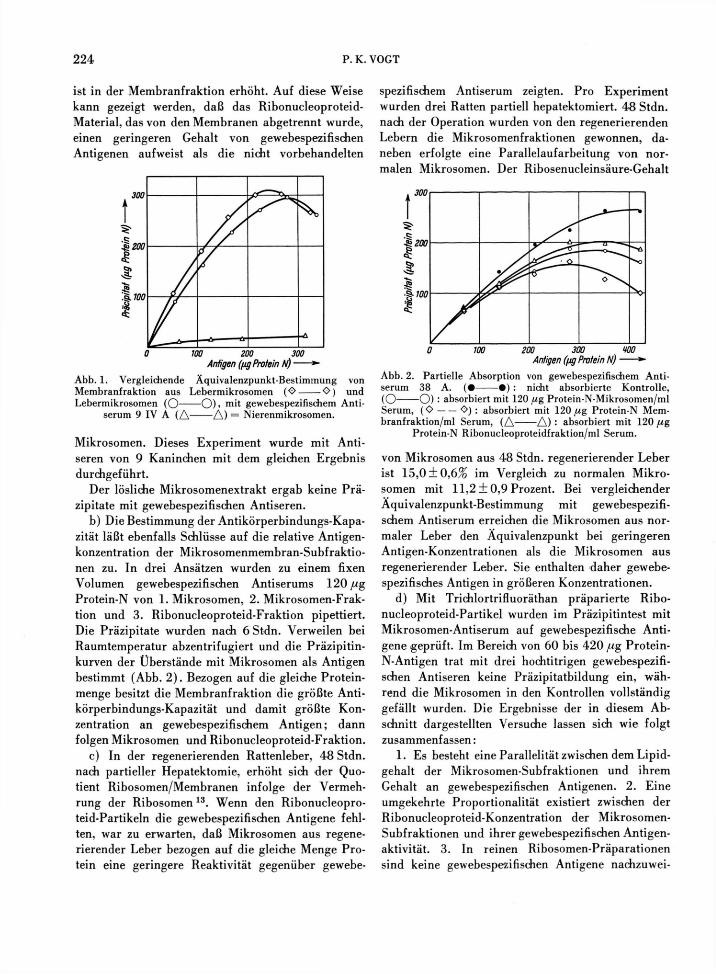
ist in der Membranfraktion erhöht. Auf diese Weise kann gezeigt werden, daß das Ribonucleoproteid-Material, das von den Membranen abgetrennt wurde, einen geringeren Gehalt von gewebespezifischen Antigenen aufweist als die nicht vorbehandelten
spezifischem Antiserum zeigten. Pro Experiment wurden drei Ratten partiell hepatektomiert. 48 Stdn. nach der Operation wurden von den regenerierenden Lebern die Mikrosomenfraktionen gewonnen, da-neben erfolgte eine Parallelaufarbeitung von nor-malen Mikrosomen. Der Ribosenucleinsäure-Gehalt
300
\ . C
200
i .§.700
0 700 200 300 Antigen (/ug Protein N)
Abb. 1. Vergleichende Äquivalenzpunkt-Bestimmung von Membranfraktion aus Lebermikrosomen (O O ) und Lebermikrosomen ( O O ) » m i t gewebespezifischem Anti-
serum 9 IV A ( A A ) = Nierenmikrosomen.
Mikrosomen. Dieses Experiment wurde mit Anti-seren von 9 Kaninchen mit dem gleichen Ergebnis durchgeführt.
Der lösliche Mikrosomenextrakt ergab keine Prä-zipitate mit gewebespezifischen Antiseren.
b) Die Bestimmung der Antikörperbindungs-Kapa-zität läßt ebenfalls Schlüsse auf die relative Antigen-konzentration der Mikrosomenmembran-Subfraktio-nen zu. In drei Ansätzen wurden zu einem fixen Volumen gewebespezifischen Antiserums 120 /ug Protein-N von 1. Mikrosomen, 2. Mikrosomen-Frak-tion und 3. Ribonucleoproteid-Fraktion pipettiert. Die Präzipitate wurden nach 6 Stdn. Verweilen bei Raumtemperatur abzentrifugiert und die Präzipitin-kurven der Überstände mit Mikrosomen als Antigen bestimmt (Abb. 2). Bezogen auf die gleiche Protein-menge besitzt die Membranfraktion die größte Anti-körperbindungs-Kapazität und damit größte Kon-zentration an gewebespezifischem Antigen; dann folgen Mikrosomen und Ribonucleoproteid-Fraktion.
c) In der regenerierenden Rattenleber, 48 Stdn. nach partieller Hepatektomie, erhöht sich der Quo-tient Ribosomen/Membranen infolge der Vermeh-rung der Ribosomen 13. Wenn den Ribonucleopro-teid-Partikeln die gewebespezifischen Antigene fehl-ten, war zu erwarten, daß Mikrosomen aus regene-rierender Leber bezogen auf die gleiche Menge Pro-tein eine geringere Reaktivität gegenüber gewebe-
300
sä 200 £ t 3s £700
0 100 200 300 100 Antigen (ng Protein N)
Abb. 2. Partielle Absorption von gewebespezifischem Anti-serum 38 A. ( • • ) : nicht absorbierte Kontrolle, ( O O ) : absorbiert mit 120 /ug Protein-N-Mikrosomen/ml Serum, ( O O) : absorbiert mit 120 [ig Protein-N Mem-branfraktion/ml Serum, ( A A ) : absorbiert mit 120 jxg
Protein-N Ribonucleoproteidfraktion/ml Serum.
von Mikrosomen aus 48 Stdn. regenerierender Leber ist 15,0 + 0,6% im Vergleich zu normalen Mikro-somen mit 11,2 ± 0,9 Prozent. Bei vergleichender Äquivalenzpunkt-Bestimmung mit gewebespezifi-schem Antiserum erreichen die Mikrosomen aus nor-maler Leber den Aquivalenzpunkt bei geringeren Antigen-Konzentrationen als die Mikrosomen aus regenerierender Leber. Sie enthalten daher gewebe-spezifisches Antigen in größeren Konzentrationen.
d) Mit Trichlortrifluoräthan präparierte Ribo-nucleoproteid-Partikel wurden im Präzipitintest mit Mikrosomen-Antiserum auf gewebespezifische Anti-gene geprüft. Im Bereich von 60 bis 420 /ug Protein-N-Antigen trat mit drei hochtitrigen gewebespezifi-schen Antiseren keine Präzipitatbildung ein, wäh-rend die Mikrosomen in den Kontrollen vollständig gefällt wurden. Die Ergebnisse der in diesem Ab-schnitt dargestellten Versuche lassen sich wie folgt zusammenfassen:
1. Es besteht eine Parallelität zwischen dem Lipid-gehalt der Mikrosomen-Subfraktionen und ihrem Gehalt an gewebespezifischen Antigenen. 2. Eine umgekehrte Proportionalität existiert zwischen der Ribonucleoproteid-Konzentration der Mikrosomen-Subfraktionen und ihrer gewebespezifischen Antigen-aktivität. 3. In reinen Ribosomen-Präparationen sind keine gewebespezifischen Antigene nachzuwei-

sen. Diese Ergebnisse sind durch die Annahme zu erklären, daß die gewebespezifischen Mikrosomen-Antigene ausschließlich in den Membranen des endo-plasmatischen Reticulums vorkommen. Die Aktivität, die sich in mit Citrat extrahierten Ribonucleoproteid-Fraktionen findet, ist auf eine Beimengung von Membranen zurückzuführen, wie sie im Elektronen-mikroskop und durch Phospholipid-Bestimmungen nachgewiesen werden kann.
Diskussion
Kombiniert mit geeigneten Fraktionierungs-Methoden vermag die immunologische Analyse von subzellulären Strukturen den Auflösungsbereich des Elektronenmikroskops zu erreichen.
Die Lokalisation der gewebespezifischen Mikro-somen-Antigene in den Membranen des endoplas-matischen Reticulums ist von besonderem Interesse im Zusammenhang mit dem Verlust solcher Anti-gene während der Carcinogenese in der Leber der Ratte und der Maus14. Elektronenmikroskopische Untersuchungen an verschiedenen Tumoren lassen keine charakteristische Veränderung des endoplasma-tischen Reticulums erkennen, die allen Krebszellen gemeinsam wäre. Am häufigsten findet sich jedoch die Degradation der Membranen 15. Für die Verän-derung der Membrankomponente des endoplasmati-schen Reticulums während der Carcinogenese in der Rattenleber sprechen auch Beobachtungen, die an Tumormikrosomen in vitro gemacht wurden. Glu-cose-6-Phosphatase, ein Enzym, das ausschließlich in den Mikrosomen-Membranen vorkommt, fehlt im Hepatom 16. Der Phospholipidgehalt der Hepatom-Mikrosomen ist reduziert17. Die Hepatomzelle ist reicher an Ribosomen als die normale Leberzelle 18. Als erste Effekte bei der Fütterung von Azofarbstof-fen fanden P O R T E R und B R U N I mit dem Elektronen-mikroskop tiefgreifende Veränderungen in der Organisation der endoplasmatischen Reticulum-Membranen. In den Primärhepatomen fehlen be-
13 J. M. PRICE u. A. K. LAIRD , Cancer Res. 1 0 , 650 [1950]; A. v. D. DECKEN u. T . HULTIN , Exp. Cell. Res. 1 4 , 88 [1958].
1 4 E . W E I L E R , Z. Naturforschg. 1 1 b, 31 [1956]; G. I. ABELJEW,
S . A . AWENIROWA , N . W . ENGELHARDT, S . L . W A I D A K O W A U.
G. I. STEPANTSCHENOK-RUDNIK, Dokl. Akad. Nauk S S S R 1 2 4 ,
1328 [1959]. 15 W. BERNHARD , Cancer Res. 18, 491 [1958]. 16 G. W E B E R U. A. CANTERO , Cancer Res. 1 5 , 105 [1955]. 1 7 G . W E B E R U. A . CANTERO , Exp. Cell Res. 1 3 , 1 2 5 [ 1 9 5 7 ] . 1 8 M . L. PETERMANN , N. A. MIZEN U. M . G. HAMILTON , Cancer
Res. 16, 620 [1956]. 19 J. C . ARCOS U. M . A. ARCOS , Biochim. biophysica Acta [Am-
sterdam] 28, 9 [1958],
stimmte Membrantypen24. Hepatom-Mikrosomen haben ihre Quellungsfähigkeit, die wahrscheinlich auf Eigenschaften der Lipoproteid-Membran zurück-geht, verloren19. Eine weitere Veränderung cyto-plasmatischer Partikel während der Carcinogenese besteht in dem Auftreten von hepatomspezifischen Antigenen, für deren Existenz neuere Versuche spre-chen 20.
Von den beiden morphologischen Bausteinen des endoplasmatischen Reticulums haben die Ribosomen bisher die größere Beachtung gefunden, wegen ihrem Vorkommen in allen daraufhin geprüften Zellen, ihrer gleichförmigen Struktur und chemischen Zu-sammensetzung und ihrer hohen metabolischen Ak-tivität in der cytoplasmatischen Proteinsynthese21. Weit weniger ist über die Funktion der Mikrosomen-Membranen und ihre Beziehungen zu den Ribo-somen bekannt. Die Lokalisation gewebespezifischer Antigene im endoplasmatischen Reticulum bringt die Membrankomponente in engen Zusammenhang mit der Differenzierung der Zelle. In der Leber-parenchymzelle wird das Cytoplasma durch die lamel-lenartigen Barrieren des endoplasmatischen Reticu-lums in ein System flacher Gefäße unterteilt, die durch Anastomosen miteinander in Verbindung ste-hen. Vermutlich können solche Strukturen Enzym-systeme und Stoffwechselprodukte in einem Status der „Semiisolierung" halten und so einen regulie-renden Einfluß auf sie ausüben 22. Ein anderer Hin-weis für die Funktion der Lipidmembranen besteht in der Beobachtung, daß für die Aktivität von DPNH-Cytochrom-c-Reduktase die Anwesenheit von bestimmten Lipiden erforderlich ist23. Dieses Enzym läßt sich durch Lipidextraktion reversibel inaktivie-ren, und die erneute Zugabe von Lipiden vermag die ursprüngliche Enzymaktivität wiederherzustel-len.
Herrn Prof. Dr. H. F R I E D R I C H - F R E K S A danke ich für mannigfache, großzügige Förderung und für sein stetes, kritisches Interesse an meiner Arbeit.
2 0 W . J. GELSTEIN , Voprosy Onkologii 4, 526 [1958]; L. A .
ZILBER, G . I . ABELJEW, S . A . AWENIROWA , N . W . ENGELHARDT
u. S . L . W A I D A K O W A , Dokl. Akad. Nauk S S S R 124, 937 [1959],
21 R. B. LOFTFIELD, Progr. Biophysics Biophysiol. Chem. 8 ,
348 [1957] ; F. H. CRICK , Symposia Soc. exp. Biol. 12, 138 [1958].
2 2 G . C. MUELLER , Cancer Res. 1 7 , 490 [1957]. 2 3 K . O . DONALDSON, A . NASON , K . B . MOORE U . R . H . GARRET,
Biochim. biophysica Acta [Amsterdam] 26, 665 [1957]. R. B. CRAWFORD, M . MORRISON U. E . STOTZ , Biochim. bio-physica Acta [Amsterdam] 33, 543 [1959].
2 4 K . R. PORTER U. C. BRUNI , Cancer Res. 1 9 , 997 [1959].


![Template for Electronic Submission to ACS Journals · Web viewS14 Figures showing the [125I]-CCL2 equilibrium displacement curves of compounds 1, 7, 8, 15a, 15b and 37a-d. S15 Figures](https://img.pdfslide.org/doc/110x75/5f9b5c625e650752db153374/template-for-electronic-submission-to-acs-journals-web-view-s14-figures-showing.jpg)





![NOTIZEN - Max Planck Societyzfn.mpdl.mpg.de/data/Reihe_B/15/ZNB-1960-15b-0323_n.pdf · luftempfindliches Nitrosyltricarbonylferrat( I)-Anion5, [Fe(CO)3NO]e, bildet, das analog den](https://img.pdfslide.org/doc/110x75/60b850de1b632226cb1aa457/notizen-max-planck-luftempfindliches-nitrosyltricarbonylferrat-i-anion5-feco3noe.jpg)




















