Embed Size (px)
Citation preview

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
1010 F. A. ANDERER
Versuche zur Bestimmung der serologisch determinanten Gruppen
des Tabakmosaikvirus
V o n F. A . A n d e r e r
Aus dem Max-Planck-Institut für Virusforschung, Tübingen(Z. Naturforschg. 18 b, 1010— 1014 [1963] ; eingegangen am 26. August 1963)
Twenty peptides of tobacco mosaic virus protein covering the whole amino acid sequence of the polypeptide chain have been tested for their capacity to inhibit the quantitative precipitation of tobacco mosaic virus with the specific antibodies. Only four amino acid sequences, representing15 — 20% of the polypeptide chain, have been found to diminish quantitative precipitation of TMV with anti TMV serum. Lack of additivity of the individual inhibition values gives evidence that the specific antibodies are not directed against isolated amino acid sequences.
Die Bestimmung der serologisch determinanten
Gruppen von Antigenen ist eines der Hauptprobleme
der Immunchemie. Untersuchungen an Polysaccha
rid-Antigenen 1 und Faserprotein-Antigenen 2 haben
gezeigt, daß hier die serologisch determinanten
Gruppen mit den terminalen Sequenzen der Poly
saccharide bzw. der Polypeptidketten identisch sind.
Bei globulären Proteinen haben entsprechende Unter
suchungen noch keine klaren Ergebnisse erbracht.
Es wird jedoch allgemein angenommen, daß bei der
serologisch determinanten Struktur von globulären
Proteinen nicht nur bestimmte Sequenzen der Pri
märstruktur maßgebend sind, sondern daß zusätzlich
durch die sekundäre und tertiäre Struktur an der
Oberfläche des Moleküls ein bestimmtes räumliches
„Muster“ ausgebildet wird, das dann für die spe
zifischen serologischen Reaktionen mitverantwortlich
ist.
Zur endgültigen Klärung dieser Frage schien das
Tabakmosaikvirus (TMV) ein besonders geeignetes
Untersuchungsobjekt, bei dem zunächst die an der
serologisch determinanten Struktur beteiligten Ami
nosäuresequenzen ermittelt werden sollten, während
die Versuche über den Einfluß der tertiären Struktur
in einer anderen Arbeit beschrieben sind 3. Bekannt
lich besteht das TMV aus einem Molekül Ribo-
1 Übersicht: E. A. K abat u. M . M . M a y e r , Experimental Im- munochemistry. C. C. Thomas Publ. 1962, Springfield, 111.
2 J. J. C e b r a , J. Immunology 86. 205 [1961].3 F. A . A n d e r e r u . D. H a n d s c h u h , Z. Naturforschg. 18 b, 1015
[1963].4 Übersicht: A. K lu g u . D. L. D. C a s p a r , Advances Virus
Res. 7,225 [I960],5 F. A . A n d e r e r , H. U h l i g , E. W e b e r u . G. S c h ra m m , Nature
[London] 186,92 [I960].6 F. A . A n d e r e r u . D. H a n d s c h u h , Z. Naturforschg. 17 b, 536
[1962].
nucleinsäure und rund 2130 gleichen Polypeptid
ketten, die im intakten Virus als strukturidentische
Proteinuntereinheiten in einer Makrohelix angeord
net sind4. Die Polypeptidketten enthalten je 158
Aminosäuren, deren Reihenfolge in früheren Arbei
ten beschrieben wurde 5~~.
Untersuchungen verschiedener Autoren8-10 wei
sen darauf hin, daß die Nucleinsäure des TMV nicht
an der serologisch determinanten Struktur beteiligt
ist. Dadurch ergibt sich die Möglichkeit, die maxi
male Größe der mit dem spezifischen Antikörper
reagierenden Oberflächeneinheit zu kalkulieren, in
dem man die maximale Antikörper-Bindungskapazi-
tät von intaktem Virus und von Virusprotein ver
gleicht. Das Verhältnis Antikörper-N/Antigen-N, das
maximal in einem Präzipitat von TMV bei extre
mem Antikörper-Uberschuß erhalten wurde, betrug
3,4 —4,1 : 1 10’ n . Bei der Verwendung von A-Pro-
tein (dissoziiertes natives Virusprotein mit Mol.-
Gew. 90 00012) als Antigen wurde dagegen ein
Verhältnis von mindestens 6 : 1 gefunden10, was
letztlich bedeutet, daß ein Antikörper mit durch
schnittlich 1 —2 Proteinuntereinheiten (Mol.-Gew.
17 530) reagiert. Diese Unterschiede sind auf steri
sche Hinderung der vollständigen Antikörper-Ab
sorption zurückzuführen, die sich beim intakten
7 A. T s u g i t a , B. T. G is h . J. Y o u n g , H. F r a e n k e l- C o n r a t , C . A. K n ig h t u . W. M. S t a n le y , Proc. nat. Acad. Sei. USA 46, 1463 [I960],
8 P. S t a r l i n g e r , Z . Naturforschg. 10 b, 339 [1955].9 H. G . A a c h , Biochim. biophysica Acta [Amsterdam] 32,
140 [1959].10 A . K le c z k o w s k i , J. Immunology 4, 130 [1961].11 G . S c h ra m m u. H. F r ie d r ic h - F r e k s a , Hoppe-Seyler’s Z. phy
siol. Chem. 270, 233 [1941].12 G . S c h ra m m , G . S c h u m a c h e r u. W. Z i l l i g , Z . Naturforschg.
10b. 481 [1955].
SEROLOGISCH DETERMINANTE GRUPPEN DES TMV 1011
Virus stärker bemerkbar macht als beim A-Protein.
Es ist daher denkbar, daß bei weiterer Dissoziation
des A-Proteins keine sterische Hinderung mehr be
steht und letztlich eine Kombination von einer Pro
teinuntereinheit mit einem Antikörper noch möglich
ist.
Diese Überlegungen weisen darauf hin, daß die
Größe der serologisch determinanten Gruppe des
TMV ungefähr dem Oberflächenanteil einer Protein
untereinheit gleichkommt, keinesfalls jedoch größer
anzunehmen ist.
Die maximale Größe einer solchen Oberflächen
einheit, die sich im intakten TMV periodisch wieder
holt, kann aus der Ganghöhe und der Zahl der
Untereinheiten je Windung der Makrohelix zu un
gefähr 700 A2 berechnet werden. Die gleiche Grö
ßenordnung wurde für den reagierenden Flächen
anteil einer Valenz eines Antikörpers (Kaninchen
serum) experimentell bestimmt13. Aus den Meß
daten der Röntgenkleinwinkelstreuung läßt sich an
dererseits der Querschnitt eines Antikörpers zu un
gefähr 800 A2 beredinen 14’ 15, was als maximaler
Wert für die reagierenden Bezirke angesehen wer
den kann, wenn man annimmt, daß diese durch die
entgegengesetzten Enden des langgestreckten Anti
körpermoleküls repräsentiert werden.
Es wurden nun Versuche unternommen, um die
Bereiche der Polypeptidketten, die an der serologisch
determinanten Oberfläche des TMV beteiligt sind,
näher zu charakterisieren. Einige vorläufige Ergeb
nisse wurden bereits mitgeteilt16. In der vorliegen
den Arbeit soll die Hemmung der Antigen-Antikör
per-Reaktion durch definierte Peptide untersucht wer
den, die durch enzymatische Spaltung des TMV-Pro-
teins gewonnen wurden.
Material und Methoden
Zu sämtlichen Untersuchungen wurde TMV vulgare verwendet. Das benützte Antiserum stammte von einem einzigen Kanindien, um individuelle Unterschiede in der Antikörper-Spezifität auszuschließen.
Das Peptidmaterial wurde durch enzymatische Spaltung von denaturiertem TMV-Protein nach früher beschriebenen Methoden erhalten 6> 17,18. Die gereinigten Peptide wurden auf Sephadex G 25-Säulen (4-160 cm)
entsalzt. Bei hoch aggregierten Peptidlösungen wurde das maximale Teilchengewicht der Aggregate in der Phywe-Ultrazentrifuge (Normalzelle) oder mit Sepha- dex-Säulen bestimmt. Die absolute Konzentration der Peptide wurde nach Totalhydrolyse mit dem automatischen Aminosäure-Analysator (Beckman-Spinco) festgestellt. Folgende Peptide wurden untersucht (in Klammern die maximale Peptidkonzentration in //Mol/ml) :
Pos. 1-10 ( 4,3) Pos. 93-112 (10,0)1-41 (10,0) 113-122 (26,6)
17-23 (10,0) 114-122 (12,0)18-23 (11,2) 123-134 (2,25) 42-46 (24,0) 123-128 (10,0) 47-61 (11,9) 129-131 ( 5,7)62 — 68 (11,5) 135-141 (38,0)69-71 ( 9,0) 142-158 (12,8)72-90 ( 9,8) 151-158 (10,0)91-92 (38,0) 153-158 (14,0)
Die Benennung der Peptide erfolgte entsprechend der Positionen der Aminosäurereste in der Polypeptidkette 6.
P r ä z i p i t a t i o n s h e m m u n g s - T est
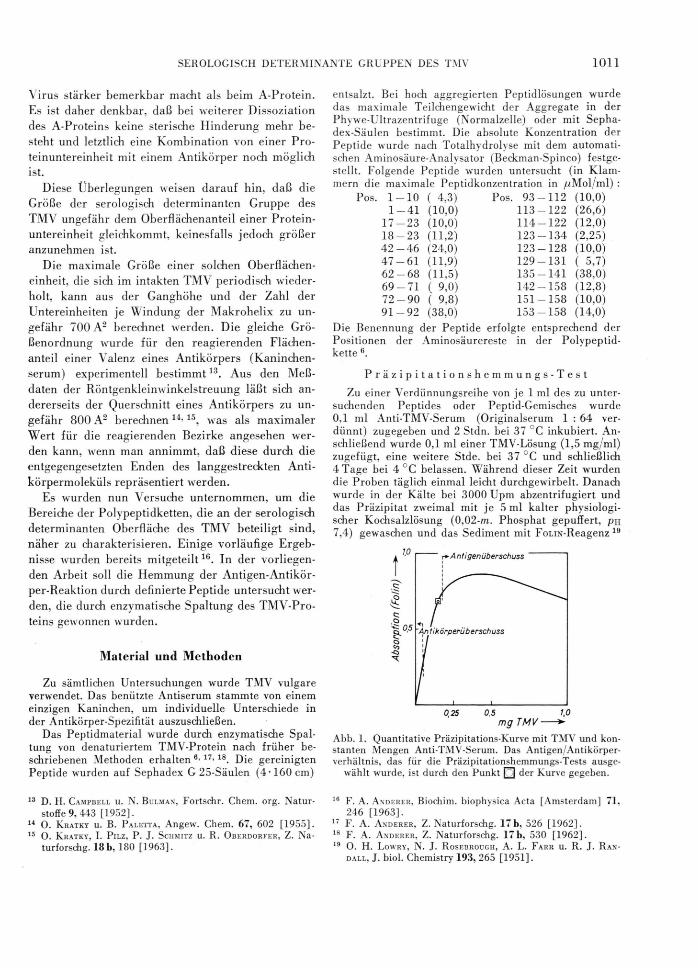
Zu einer Verdünnungsreihe von je 1 ml des zu untersuchenden Peptides oder Peptid-Gemisches wurde0,1 ml Anti-TMV-Serum (Originalserum 1 : 64 verdünnt) zugegeben und 2 Stdn. bei 37 °C inkubiert. Anschließend wurde 0,1 ml einer TMV-Lösung (1,5 mg/ml) zugefügt, eine weitere Stde. bei 37 °C und schließlich 4 Tage bei 4 °C belassen. Während dieser Zeit wurden die Proben täglich einmal leicht durchgewirbelt. Danach wurde in der Kälte bei 3000 Upm abzentrifugiert und das Präzipitat zweimal mit je 5 ml kalter physiologischer Kochsalzlösung (0,02-m. Phosphat gepuffert, ph 7,4) gewaschen und das Sediment mit FoLiN-Reagenz 19
mg TMV ■
Abb. 1. Quantitative Präzipitations-Kurve mit TMV und konstanten Mengen Anti-TMV-Serum. Das Antigen/Antikörperverhältnis, das für die Präzipitationshemmungs-Tests ausge
wählt wurde, ist durch den Punkt □ der Kurve gegeben.
13 D. H. C a m p b e l l u . N. B u lm a n , Fortschr. Chem. org. Naturstoffe 9, 443 [1952],
14 0. K r a t k y u . B. P a l e t t a , Angew. Chem. 67, 602 [1955].15 O . K r a t k y , I. P i l z , P . J. S c h m itz u . R. O b e r d ö r f e r , Z. Na
turforschg. 18 b, 180 [1963].
16 F. A. A n d e r e r , Biochim. biophysica Acta [Amsterdam] 71, 246 [1963].
17 F. A . A n d e r e r , Z. Naturforschg. 17 b, 526 [1962].18 F. A . A n d e r e r , Z. Naturforschg. 17 b, 530 [1962].19 0. H. L o w r y , N. J. R o s e b r o u g h , A. L. F a r r u . R . J. R a n
d a l l , J. biol. Chemistry 193, 265 [1951].
1012 F. A. ANDERER
quantitativ bestimmt. Mit den Kontrollen (statt 1,0 ml Peptidlösung, 1,0 ml physiologische Kochsalz-Phosphat- Lösung) wurde analog verfahren.
Für diese Hemmungsteste und für die Kontrollen wurde die Konzentration von TMV- und Anti-TMV- Serum so gewählt, daß die Bedingungen einem Punkt innerhalb der Äquivalenzzone entsprachen, der jedoch ziemlich nahe an der Grenze zum Gebiet des Antigen- überschusses lag (s. Abb. 1). Damit sollte erreicht werden, daß möglichst alle präzipitierenden Antikörper divalent mit dem Antigen verbunden sind, um die Hemmung der Präzipitinreaktion durch die Peptid-Haptene empfindlicher zu machen. Der Anteil der Antikörper im Präzipitat der Kontrollen betrug ungefähr 25 Prozent.
Ergebnisse
Partielle enzymatische Hydrolysate von denatu
riertem TMV-Protein, die durch Einwirkung von
Trypsin, Chymotrypsin, Pepsin oder Papain erhal
ten wurden, zeigten im Präzipitationshemmungs-
Test keinerlei Hemmeffekt. Der antigene Charakter
des TMV-Proteins ist verschwunden, wenn maximal
4 Peptidbindungen durch Einwirkung der einzelnen
Enzyme gespalten sind 3. Die in Lösung vorhande
nen, noch relativ großen Spaltpeptide hatten auch
keinerlei Hemmwirkung auf die Präzipitin-Reaktion
von TMV und Anti-TMV-Serum.
Der Hemmungseffekt von Haptenen ist jedoch
stark von der Konzentration abhängig. Die maxi
male Konzentration an gespaltenen Polypeptidket
ten, die in den enzymatischen Hydrolysaten erreicht
werden konnte, betrug ungefähr 0.5 //Mol pro ml.
Das entspricht einem 60-fachen Überschuß gegen
über der Menge der Peptide, die bei den Hemmungs
versuchen gebunden im Virusantigen Vorlagen. Die
ser Überschuß ist möglicherweise nicht groß genug,
um einen meßbaren Hemmungseffekt zu zeigen. Es
war daher angebracht, einzelne gereinigte Peptide
oder Gemische aus zwei bis drei Peptiden auf ihren
Haptencharakter zu untersuchen. Durch geeignete
Auswahl der Antigen- und Antiserum-Konzentration
konnte schließlich das Antigen/Hapten-Verhältnis
1 : 1000 bis 1 : 4000 erhalten werden. Dabei stellte
sich heraus, daß mehrere Spaltstücke aus der Poly
peptidkette des TMV einen deutlichen Hemmungs
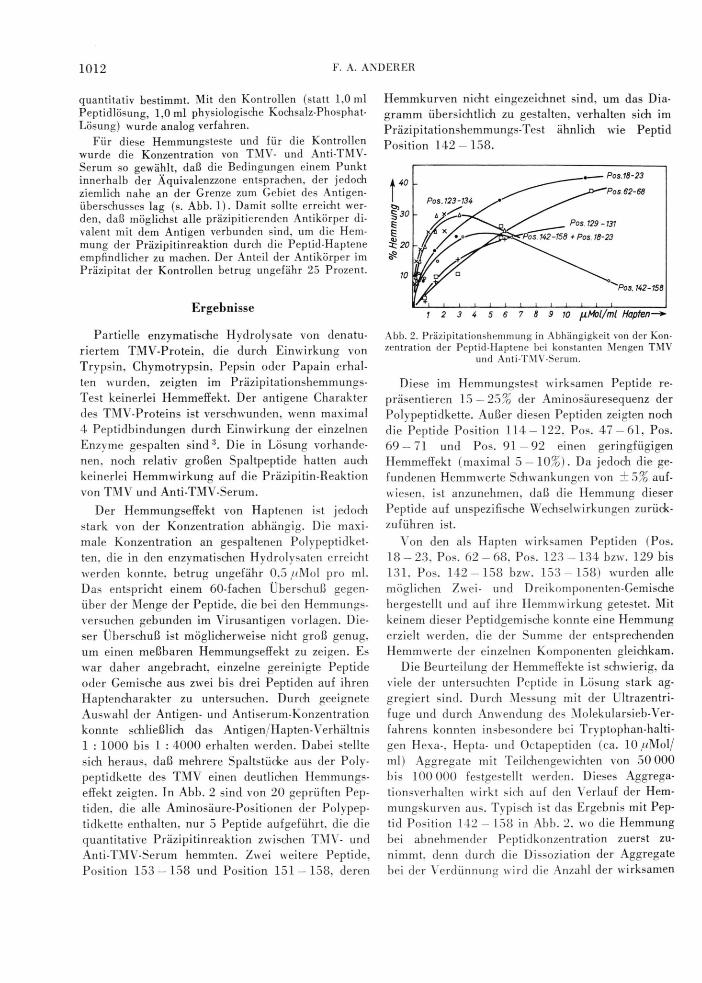
effekt zeigten. In Abb. 2 sind von 20 geprüften Pep
tiden, die alle Aminosäure-Positionen der Polypep
tidkette enthalten, nur 5 Peptide aufgeführt, die die
quantitative Präzipitinreaktion zwischen TMV- und
Anti-TMV-Serum hemmten. Zwei weitere Peptide,
Position 153 — 158 und Position 151 — 158, deren
Hemmkurven nicht eingezeichnet sind, um das Dia
gramm übersichtlich zu gestalten, verhalten sich im
Präzipitationshemmungs-Test ähnlich wie Peptid
Position 142 — 158.
Abb. 2. Präzipitationshemmung in Abhängigkeit von der Konzentration der Peptid-Haptene bei konstanten Mengen TMV
und Anti-TMV-Serum.
Diese im Hemmungstest wirksamen Peptide re
präsentieren 15 — 25% der Aminosäuresequenz der
Polypeptidkette. Außer diesen Peptiden zeigten noch
die Peptide Position 114 — 122, Pos. 47 — 61, Pos.
69 — 71 und Pos. 91—92 einen geringfügigen
Hemmeffekt (maximal 5 — 10%). Da jedoch die ge
fundenen Hemmwerte Schwankungen von ± 5% auf
wiesen, ist anzunehmen, daß die Hemmung dieser
Peptide auf unspezifische Wechselwirkungen zurück
zuführen ist.
Von den als Hapten wirksamen Peptiden (Pos.
18 - 23, Pos. 62 - 68. Pos. 123 - 134 bzw. 129 bis
131, Pos. 142 — 158 bzw. 153 — 158) wurden alle
möglichen Zwei- und Dreikomponenten-Gemische
hergestellt und auf ihre Hemmwirkung getestet. Mit
keinem dieser Peptidgemische konnte eine Hemmung
erzielt werden, die der Summe der entsprechenden
Hemmwerte der einzelnen Komponenten gleichkam.
Die Beurteilung der Hemmeffekte ist schwierig, da
viele der untersuchten Peptide in Lösung stark ag
gregiert sind. Durch Messung mit der Ultrazentri
fuge und durch Anwendung des Molekularsieb-Ver-
fahrens konnten insbesondere bei Tryptophan-halti
gen Hexa-, Hepta- und Octapeptiden (ca. 10 «Mol/
ml) Aggregate mit Teilchengewichten von 50 000
bis 100 000 festgestellt werden. Dieses Aggrega
tionsverhalten wirkt sich auf den Verlauf der Hem
mungskurven aus. Typisch ist das Ergebnis mit Pep
tid Position 142 — 158 in Abb. 2, wo die Hemmung
bei abnehmender Peptidkonzentration zuerst zu
nimmt. denn durch die Dissoziation der Aggregate
bei der Verdünnung wird die Anzahl der wirksamen

SEROLOGISCH DETERMINANTE GRUPPEN DES TMV 1013
Haptenmoleküle vergrößert. Andere, teilweise auch
unwirksame Peptide (Peptid Position 1 —41, Pos.
1 — 10 und Pos. 17 — 23) sind in konzentrierten Lö
sungen hoch aggregiert; Peptid Pos. 1 —41 bis zu
einem Teilchengewicht von über 100 000 20.
Diskussion
Einen starken Hemmeffekt zeigten in dem durch
geführten Test nur die Aminosäure-Sequenzen Pos.
18-23 , Pos. 62 - 68 , Pos. 123-134 bzw. 129
bis 131 und Pos. 142 — 158 bzw. 153 — 158. Eine
Beteiligung aller hemmenden Peptide an der serolo
gisch determinanten Gruppe bzw. am Oberflächen
anteil der Proteinuntereinheit im TMV ist jedoch
aus sterischen Gründen nicht möglich, da im Modell
einerseits die zur Verfügung stehende Oberfläche
nicht ausreicht und andererseits (ohne die zur Ver
fügung stehende Oberfläche zu berücksichtigen) die
Masse der Polypeptidkette an der Oberfläche des
TMV so dicht zusammengedrängt werden müßte,
daß es der ziemlich gleichmäßigen radialen Vertei
lung der Elektronendichte im intakten Virus wider
sprechen würde 22’ 23.
Sicher ist jedoch, daß die C-terminale Sequenz
der Polypeptidkette an der serologisch determinan
ten Struktur des TMV beteiligt ist. Durch Unter
suchung von H a r r i s und K n i g h t 21 ist bekannt, daß
die C-terminale Aminosäure Threonin an der Ober
fläche des TMV liegt, da sie sich durch Carboxypep-
tidase abspalten läßt. Das so behandelte Virus und
sein spezifisches Antiserum geben eine serologische
Kreuzreaktion mit intaktem TMV bzw. dem entspre
chenden Antiserum. Danach ist eine Beteiligung der
C-terminalen Aminosäure Threonin an der serolo
gisch determinanten Gruppe des TMV ziemlich wahr
scheinlich. Eine Bestätigung hierfür erhalten wir
durch die ausgeprägte Hemmwirkung der C-termina
len Peptide, Pos. 142 — 158, Pos. 151 — 158 und Pos.
153 — 158. Ferner konnte in einer früheren Arbeit16 gezeigt werden, daß ein synthetisches Antigen mit
dem C-terminalen Hexapeptid der Polypeptidkette
20 H . F r a e n k e l- C o n r a t u. L. K . R a m a c h a n d r a n , Advances Protein Chem. 14, 175 [1959].
21 J. H . H a r r i s u. C. A. K n ig h t , Nature [London] 170, 613 [1952] ; J. biol. Chemistry 214, 215 [1955].
22 D. L. D. C a s p a r , Nature [London] 177, 928 [1956].23 R. E. F r a n k l i n , A. K lu g u . H . C. H o lm e s (1957), Ciba
Foundation Symp. on the Nature of Viruses, p. 39.Churchill, London.
des TMV als determinanter Gruppe mit TMV serolo
gisch verwandt ist, da die entsprechenden Antikörper
mit TMV präzipitieren und die Virus-Infektiosität
neutralisieren.
Auch für die Sequenz Pos. 18 — 23 gibt es Be
funde, die auf eine exponierte Lage an der Virus
oberfläche hinweisen und dadurch eine Beteiligung
an der serologisch determinanten Struktur wahr
scheinlich machen. Bei der Behandlung des Virus
mit heißer Trichloressigsäure24 oder Hydroxyl
amin 2o- 26 wird bevorzugt eine einzige labile Asp-
Pro-Bindung nahezu quantitativ gespalten, obwohl
insgesamt zwei Asp-Pro-Bindungen in der Polypep
tidkette des TMV Vorkommen (Pos. 19 — 20 und
77 — 78 )27. Die Befunde von Braunitzer 28 weisen
darauf hin, daß die Peptidbindung zwischen Posi
tion 19 und 20 bevorzugt gespalten wird, was durch
eine sterisch exponierte Lage an der Oberfläche des
Virus erklärt werden könnte.
Für die anderen beiden als Hapten wirksamen
Aminosäuresequenzen (Pos. 62 — 68 und 123 — 134)
gibt es vorläufig keine weiteren Kriterien für ihre
räumliche Anordnung im nativen Virus. Die Hemm-
kurven allein können hierbei nicht als entscheidendes
Kriterium benützt werden, da einerseits die absolute
Konzentrationen der Peptidhaptene wegen der Ag
gregatbildung in Lösung nicht der Zahl der wirk
samen Haptenmoleküle entspricht und andererseits
die Hemmwerte der einzelnen Peptide wegen ver
schieden starker Aggregation nicht zueinander in Be
ziehung gesetzt werden können.
Im Vergleich mit analogen Untersuchungen an
Polysacchariden und Faserproteinen hat der Präzipi
tationshemmungs-Test mit Haptenen im System
TMV/Anti-TMV-Serum nicht die Erwartungen er
füllt. Die maximale Hemmwirkung, die mit den ge
reinigten Peptidfraktionen erreicht wurde, ist we
sentlich niedriger als die maximale Hemmwirkung
der serologisch determinanten terminalen Oligo-Sac-
charide im Dextran/Antidextran-System von Kabat 29 oder der terminalen Peptide im System Seiden
fibroin/An tiseidenfibroin von Cebra 2.
24 G. S c h ra m m u . G. B r a u n i t z e r , Z. Naturforschg. 8 b, 61[1953].
25 G. B r a u n i t z e r , Biochim. biophysica Acta [Amsterdam] 19,574 [1956],
26 L. K . R a m a c h a n d r a n u . K . N a r i t a , Biochim. biophysicaActa [Amsterdam] 30,616 [1958].
27 F. A . A n d e r e r , Z. Naturforschg. 14 b, 363 [1959].28 G. B r a u n i t z e r , Naturwissenschaften 42, 371 [1955].29 E. A. K a b a t , J. Amer. chem. Soc. 76, 3709 [1954].

1014 SEROLOGISCH DETERMINANTE GRUPPEN DES TMV
Diesem Befund können zwei Ursachen zugrunde
liegen. Das spezifische Serum gegen TMV besteht
aus einem Gemisch von Antikörpern, die jeweils ge
gen einzelne Aminosäure-Sequenzen an der Ober
fläche des Virus gerichtet sind, so daß immer nur
ein bestimmter Anteil der Antikörper durch ein ein
zelnes Peptidhapten quantitativ gehemmt werden
kann, während die verbliebenen Antikörper mit
TMV präzipitieren. Dagegen spricht jedoch, daß sich
die Hemmwerte der einzelnen Peptide nicht additiv
verhalten, wenn zwei oder drei der wirksamen Pep
tide im Gemisch als Hapten eingesetzt werden. Eine
andere Erklärung bietet sich an. wenn man annimmt,
daß die serologisch determinanten Gruppen des
TMV nicht durch einzelne Sequenzbereiche der Pri
märstruktur gebildet werden, sondern durch mehrere
Sequenzbereiche, die in der Primärstruktur durch
andere Aminosäure-Sequenzen getrennt sind, jedoch
durch entsprechende Faltung der Polypeptidkette im
nativen Zustand räumlich eng an der Oberfläche des
Virus zusammenrücken und dadurch eine größere
Struktureinheit bilden. Der spezifisch affine Mole
külbezirk im Antikörper müßte daher entsprechend
groß sein, was letztlich auch die Wahrscheinlichkeit
für eine unspezifische Wechselwirkung zwischen
Testpeptiden und Antikörper vergrößert. Anderer
seits erscheint dadurch auch die relativ kleine Hemm
wirkung der in Abb. 2 angeführten Peptide ver
ständlich. da diese gemäß dem obigen Postulat nur
als partiell determinante Gruppen anzusehen sind.
Zur endgültigen Klärung dieser Frage müßten je
doch weitere Kriterien, insbesondere die tertiäre
Struktur betreffend, herangezogen werden, mit
denen sich die nächste Arbeit befaßt 3.
Zusammenfassend ist festzustellen, daß unter ge
wissen Vorbehalten die Präzipitationshemmungs-
Reaktion auch bei globulären Proteinen zur Bestim
mung der serologisch determinanten Aminosäure-
Sequenz benützt werden kann. In der vorliegenden
Arbeit konnte auf Grund der durchgeführten Unter
suchung für einen Großteil der Peptide aus TMV-
Protein (75 — 85% der Aminosäure-Sequenz) ausge
schlossen werden, daß sie an den serologisch deter
minanten Gruppen des TMV beteiligt sind. Für die
endgültige Festlegung der entsprechenden Amino
säure-Sequenzen müssen jedoch noch andere Krite
rien herangezogen werden, wie z. B. die serologische
Untersuchung von künstlichen Antigenen mit den in
Frage kommenden Peptiden als determinante Grup
pen, was im Falle des C-terminalen Hexapeptids aus
TMV-Protein zu einem positiven Ergebnis geführt
hat16.
Prof. Dr. G. S c h ra m m danke ich für die Förderung der Arbeit und Frl. H. v o n S c h ü t z für ihre sorgfältige technische Assistenz.





























