Embed Size (px)
Citation preview

Originalarbeiten . Original Papers
Fachbereich Biologie - Botanisches Institut -Philipps-Universitat Marburg/Lahn
Physiologische Unterschiede im Jodstoffwechsel von Asparagopsis und Falkenbergia
Physiological Variation in the Iodine Metabolism of Asparagopsis and F alkenbergia
JOHANNA KNAPPE und DIETRICH WERNER
Mit 5 Abbildungen
Eingegangen am 5. September 1974
Summary
1. Asparagopsis armata and Falkenbergia ru/olanosa, representing different generations of the same species of red algae, take up iodine from the medium with significantly different rates. The former is also a more efficient scavenger by a factor of ten (2 X 10-3 pg at III remaining in the medium for Asparagopsts, 2 X 10-2 pg at 1/1 for Falkenbergia).
2. The uptake of iodine is independent from light in both forms. Inhibitors of photosynthesis do not influence the uptake of iodine.
3. About 90 DID of the incorporated iodine can be extracted with 1 n HCl or with water at 70°C from Asparagopsis versus only 50 Ofo from Falkenbergia. By t-butanol almost 8 times as much labelled iodine can be extracted from Asparagopsis. However the same amount of iodine is extracted from both forms by 3 DID ammonia in methanol.
4. Treatment with trypsin or pronase solubilize additionally from both forms the same amount of 131 J, about 20 DID of the total activity
5. Two-dimensional thin-layer chromatography of extracts with 3 Ofo ammonia in methanol reveal, that more than 99 % of the 1311 activity from Asparagopsis is concentrated on one spot, whereas the 1311 activity from Falkenbergia is separated in a ratio of 42 : 40 : 15 : 3 on four different spots with very different rl-values. The chromatography of water extracts produced comparable differences in the two forms.
6. The results are compared with the physiological data available in the literature on iodine metabolism in red and brown algae.
Zusammenfassung
Bei den beiden im Generationswechsel stehenden Rotalgenformen Asparagopsis und Falkenbergia werden im Jodstoffwechsel signifikante Unterschiede hinsichtlich der Aufnahmerate, der im Medium verbleibenden Endkonzentrationen an Jod und der Extrahierbarkeit jodhaltiger Komponenten mit verschiedenen Uisungsmitteln nachgewiesen. Diinnschichtchromatogra-
z. P/lanzenphysiol. Bd. 75. S. 277-286. 1975.

278 J. KNAPPE und D. WERNER
phische Trennungen zeigen eine wesentlich verschiedene Zusammensetzung jodhaltiger Metabolite bei beiden Formen.
Einleitung
Spurenelemente spiel en im Stoffwechsel der Pflanzen eine wichtige Rolle. Mangelzustande konnen sich so stark auswirken, dag der Einflug auf die normale Morphologie und die Differenzierung direkt sichtbar wird. Jod ist das Element mit dem hochsten Molekulargewicht, das fUr viele Organismen des Tier- und Pflanzenreichs als Spurenelement essentiell ist. Wahrend die Funktion des Jods fur den Schilddrusenstoffwechsel nach verschiedenen Gesichtspunkten erforscht wurde, ist seine Bedeutung fur die Pflanzen noch weitgehend unbekannt. Meeresalgen nehmen Jod aus dem Meerwasser auf. Insbesondere sind viele Braunalgen reich an Jod, bei Laminaria z. B. betragt der Jodgehalt bis zu 1 Ofo des Trockengewichtes (SHAW, 1959). Man verwendet daher Laminaria-Arten zur technischen Jodgewinnung. Auch Rotalgen reichern Jod an. Einige Gattungen differenzieren fiir die Speicherung besondere Zellen, die von KYLIN (1915) als Blasenzellen bezeichnet wurden. Die Bedeutung der Blasenzellen ist unklar; Jod konnte auch in anderen Zellen des Algenthallus nachgewiesen werden.
Die beiden Rotalgen Asparagopsis und Falkenbergia aus der Familie der Bonnemaisoniaceae stehen miteinander in Generationswechsel. Dabei stellt Asparagopsis, wie man erst aus Kulturversuchen weig (J. und G. FELDMANN, 1942), den Gametophyten dar, wahrend Falkenbergia der dazugehorige Tetraporophyt ist. Aus den morphologisch-anatomischen Unterschieden der Thalli erklaren sich die urspriinglichen Gattungsbezeichnungen. Halt man diese Algen in Kultur, so benotigen sie uber die im naturlichen Meerwasser enthaltenen Jodmenge (ca. 3,9 X 10-1 fig at J/1) hinaus noch eine weitere Jodzugabe. Lagt man diese weg, so zeigen die Algen reversible morphologische Veranderungen im Thallusaufbau, wie VON STOSCH (1963) zeigen konnte. Der Jodmangel bewirkt eine charakteristische Veranderung der Thallusdifferenzierung, die sich beim Gametophyten Asparagopsis in einer Reduzierung der Seitenaste und in einem insgesamt sukkulenten Kriippelwachstum augert (v. STOSCH, 1963). Das Wachstum des Sporophyten Falkenbergia, dessen Thallus gegenuber Asparagopsis einen vereinfachten Bauplan zeigt, wird durch Jodmangel nicht so deutlich beeinflugt. Durch Zugabe jodhaltiger Verbindungen konnen diese Mangelerscheinungen bereits nach wenigen Tagen riickgangig gemacht werden. Bei der zu der Familie der Rhodomelaceae gehorenden Polysiphonia urceolata konnte eine lineare Beziehung zwischen dem Jodgehalt des Mediums und der Zunahme des Trockengewichts der Alge festgestellt werden (FRIES, 1966). Auch bei Braunalgen konnte die Notwendigkeit von Jod fiir Wachstum und Entwicklung nachgewiesen werden, z. B. fur Ectocarpus siliculosus (WOOLERY und LEWIN, 1973) und fUr Petalonia fascia (HSIAO, 1969).
Material und Methoden
Asparagopsis armata und Falkenbergia rufolanosa stammen aus der Kulturensammlung von v. STOSCH. Sie wurden vegetativ vermehrt in modifizierter Schreiber-Uisung (v. STOSCH,
Z. Pflanzenphysiol. Bd. 75. S. 277-286. 1975.

)odstoffwechsel von Asparagopsis und Falkenbergia 279
1963). In 1000 ml filtriertem Seewasser sind zusatzlich enthalten : 42,5 mg NaN03; 10,75 mg Na2HP04 A 12 H 20; 278 fig FeS04 A 7 H20; 19,8 ,ug MnClz X 4 H20 . Diese Losung wurde sterilisiert und vor Gebrauch mit folgenden Zusatzen versehen:
0,7 .ug Vitamin B12 ; 19,6 ,ug AS~03 ; 3,72 mg Na2EDTA; 166 ,ug K) fur Falkenbergia bzw. 830 pg K) fur Asparagopsis.
Die Kulturen wuchsen unter folgenden Standardbedingungen: 15 °C, 400 Lux (Asparagop sis) bzw. 200 Lux (Falkenbergia) in einem Licht-Dunkel-Wechsel von 14 : 10 Std.
131) bezogen wir als Natrium}odid in schwach alkalischer Thiosulfatlosung (pH 8-10) mit einer Ausgangsaktivitat von 1 mCi/O,1 ,llg NaJ. Die Halbwertszeit betragt 8 Tage. Fur die Versuche kam jeweils eine Pflanze in eine Petrischale mit frischer Nahrlosung ohne K), zu der in der Regel 50 ,uCi 131)/10 ml hinzugefugt wurde. Seewasser enthalt 3,9 X 10-1 fIg at ) / 1, 50 pCi )/10 ml entsprechen 3,9 X 10-3 /lg at ) / 1; 10 ml Nahrlosung enthalten 3,94 X 10-3 ,ug at )od, bei zusatzlicher anorganischer )odzugabe erhoht sich die )odkonzentration auf 5,39 X 10-2 fIg at ) 110 m!.
Zur Kontrolle der Aufnahmekinetik von 131J wurden jeweils 0,1 ml aus der Nahrlosung entnommen und 5 ml Dioxanszintillator folgender Zusammensetzung zugefugt: in 1000 ml Dioxan waren 4 g DPO, 75 mg POPOP und 100 g Naphthalin gelost.
Am Ende des Versuches wurden die Pflanzen dreimal mit 3,5 0/oiger NaCl-Losung gewaschen und anschlieBend gefricrgetrocknet. Die Trockengewichte wurden auf einer Mikroanalysenwaage bestimmt und als BezugsgroBe verwendet. Zur Bestimmung der Gesamtaktivitat in den Algen wurden diese in den Szintillationsfiaschchen mit je 1 ml «Digestifl» (Merck) uber Nacht bei 40 °C verdaut. Nach Abkuhlung kam 10 ml DioxanszintiIIator hinzu. Fur die Extraktionen wurden abgewogene Algenmengen in Zentrifugenglaser gebracht, zu jeder Probe 5 ml Extraktionsmittel (s . Tabelle 1) hinzugegeben und zu verschiedenen Zeiten jeweils 0,5 ml yom Oberstand zur Aktivitatsmessung entnommen. Zur Bestimmung der Aktivirat im Ruckstand wurde dieser zweimal mit 3,5 0/oiger NaCl gewaschen und eben falls mit «Digestin» verdaut.
Fur die Hydrolyse mit Hilfe von Pronase und Trypsin wurde jede Probe 10 sec zermorsert, mit 5 ml Trispuffer (0,05 M; pH 7,6) in ein Zentrifugenglas gespult und nach 10 min abzentrifugiert (1400 X g/5 min). Zu den Ruckstanden kamen jeweils 4 ml Enzymlosung (2 mg Enzym/ml Trispuffer). Nach Inkubation wurde abzentrifugiert (1400 X g/2 min) und die Aktivitat in aliquots des Oberstandes bestimmt.
Die Proben wurden im Tri-Carb-Fliissig-Szintillationsspektrometer (Packard, Modell 3380) bei automatischem Probenwechsel je zweimal gemessen. Die Quenchkorrekturen wurden mit der Kanalverhaltnismethode des externen Standards durchgefuhrt.
Die gemessencn Counts wurden auf mg Trockengewicht bezogen und die Gesamtaktivitat = 100 Ol D gesetzt. So konnten, unabhangig von der ZerfalIsrate, die Werte aus verschiedenen Versuchen gemittelt werden.
Fur die dunnschichtchromatographischen Trennungen wurden die Proben 5 min im Extraktionsmittel zermorsert, abzentrifugien und jeweils 0,05 ml des Oberstandes auf eine CelluloseDC-Fertigplatte (Fa. Merck) aufgetragen. Nach zweidimensionaler Entwicklung mit t-Butano1l3 Ufo NH3 (3 : 1) und s-Butanol/Eisessig/H20 (4 : 4 : 1) wurde die Lage der Flecken autoradiographisch bestimmt. Nach Festlegung der Flecken auf der Diinnschichtplatte wurde das Tragermaterial von den einzelnen Flecken mit Hilfe cines Spatels in Szintillationsflaschchen iiberfuhrt und die Aktivitat quantitativ gemessen.
Ergebnisse und Diskussion
Gibt man zu einer Asparagopsis-Kultur 50 ,LtCi 131J (Gesamtkonzentration ca. 4 X 10-1 ,Ltg at Jod/1) und verfolgt die Aufnahme, so sieht man, daB bereits nam 12 Std. 50 Ofo der Jodmenge aus der Nahrl6sung versmwunden ist. Nam einem Tag
Z. P/lanzenphysiol. Bd. 75. S. 277-286. 1975.
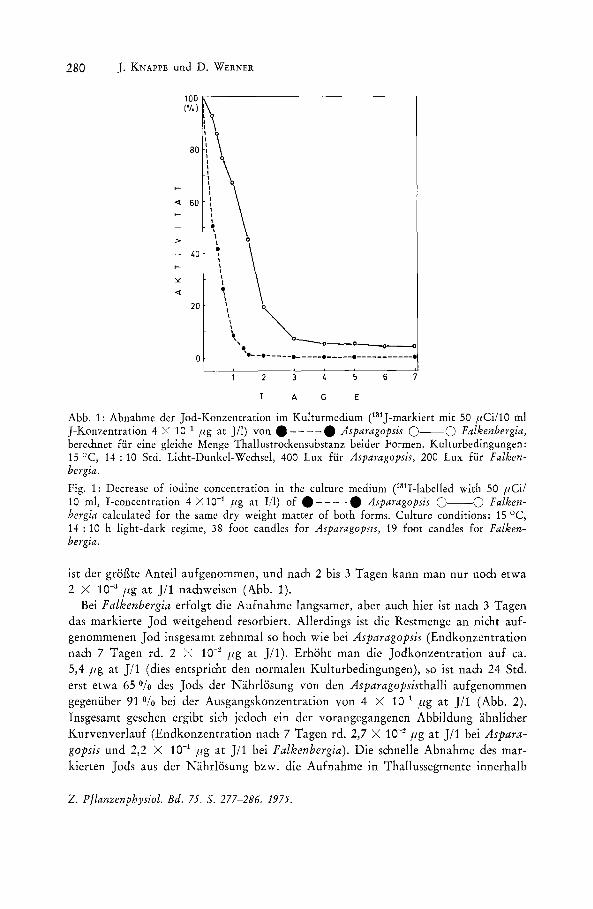
280 ]. KNAPPE und D. WERNER
100.------------------------------, ('10) I
I 0
I \ I 0
80 \ \ I 0
\\ <;. \ \
~ '" \ \ 20 \ o~o
'., ---0--0_---0 o e--e-----e _____ e _____ e ___________ •
5 6
A G E
Abb. 1: Abnahme der Jod-Konzentration im Kulturmedium (131J-markiert mit 50 ,uCi/l0 ml J-Konzentration 4 X 10·'1 fig at J/I) von .----. Asparagopsis 0--0 Falkenbergia, berechnet fur eine gleiche Menge Thallustrod,ensubstanz beider Formen. Kulturbedingungen: 15°C, 14 : 10 Std. Licht-Dunkel-Wechsel, 400 Lux fur Asparagopsis, 200 Lux fur Falkenbergia.
Fig. 1: Decrease of iodine concentration in the culture medium CS1I-labelled with 50 ,uCi/ 10 ml, I-concentration 4 X 10-1 pg at Ill) of • - - - -. Asparagopsis 0--0 Falkenbergia calculated for the same dry weight matter of both forms. Culture conditions: 15°C, 14: 10 h light-dark regime, 38 foot candles for Asparagopsls, 19 foot candles for Falkenbergia.
ist der groBte Anteil aufgenommen, und nach 2 bis 3 Tagen kann man nur noch etwa 2 X 10-s fig at J/1 nachweisen (Abb. 1).
Bei Falkenbergia erfolgt die Aufnahme langsamer, aber auch hier ist nach 3 Tagen das markierte Jod weitgehend resorbiert. Allerdings ist die Restmenge an nicht aufgenommenen Jod insgesamt zehnmal so hoch wie bei Asparagopsis (Endkonzentration nach 7 Tagen rd. 2 >; 10-2 fig at J/1). Erhoht man die Jodkonzentration auf ca. 5,4 fig at J/1 (dies entspricht den normalen Kulturbedingungen), so ist nach 24 Std. erst etwa 65 Ofo des Jods der Nahrlosung von den Asparagopsisthalli aufgenommen gegenliber 91 Ofo bei der Ausgangskonzentration von 4 X 10-1 fig at J/1 (Abb. 2). Insgesamt gesehen ergibt sich jedoch ein der vorangegangenen Abbildung ahnlicher Kurvenverlauf (Endkonzentration nach 7 Tagen rd. 2,7 X 10-2 flg at J/1 bei Asparagopsis und 2,2 X 10-1 fig at J/1 bei Falkenbergia). Die schnelle Abnahme des markierten Jods aus der Nahrlosung bzw. die Aufnahme in Thallussegmente innerhalb
Z. P/lanzenphysiol. Bd. 75. S. 277-286. 1975.
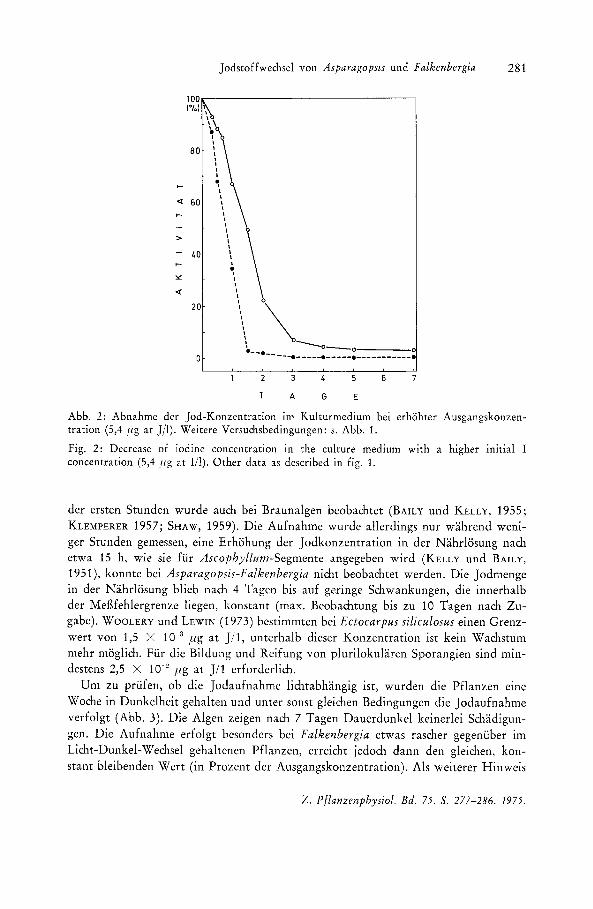
Jodstoffwechsel von Asparagopm und Falkenbergia 281
100r------------------------------, ('/,)1\:-.
I"~\ ~\
00 \\
HO \\
>
- 40
« 20
\ \ ~ I I I I
\ 0\ I ----0 __ • __ • _____ • _____ • _____ ~'-------------------·.01
6
A G E
Abb. 2: Abnahme der Jod-Konzentration im Kulturmedium bei erhohter Ausgangskonzentration (5,4 ,fig at J/I). Weitere Versuchsbedingungen: s. Abb. 1.
Fig. 2: Decrease of iodine concentration in the culture medium with a higher initial I concentration (5,4 ,llg at III). Other data as described in fig. 1.
der ersten Stun den wurde auch bei Braunalgen beobachtet (BAILY und KELLY, 1955; KLEMPERER 1957; SHAW, 1959) . Die Aufnahme wurde allerdings nur wahrend weniger Stunden gemessen, eine Erhohung der Jodkonzentration in der Nahrlosung nach etwa 15 h, wie sie fur Ascophyllum-Segmcnte angegeben wird (KELLY und BAILY, 1951), konnte bei Asparagopsis-Falkenbergia nicht beobachtet werden. Die Jodmenge in der Nahrlosung blieb nach 4 Tagen bis auf geringe Schwankllngen, die innerhalb der MefHehlergrenze liegen, konstant (max. Beobachtung bis Zll 10 Tagen nach Zugabe). WOOLERY und LEWIN (1973) bestimmten bei Ectocarpus siliculosus einen Grenzwert von 1,5 X 10-3 pg at J / 1, unterhalb dieser Konzentration ist kein Wachstum mehr moglich. Fur die Bildung und Reifung von plurilokularen Sporangien sind mindestens 2,5 X 10-" fig at J / 1 erforderlich.
Urn zu prufen, ob die Jodaufnahme lichtabhangig ist, wurden die Pflanzen eine Woche in Dunkelheit gehalten und unter sonst gleichen Bedingungen die Jodaufnahme verfolgt (Abb. 3). Die Algen zeigen nach 7 Tagen Dallerdunkel keinerlei Schadigungen. Die Aufnahme erfolgt besonders bei F alkenbergia etwas rascher gegeniiber im Licht-Dunkel-Wechsel gehaltcnen Pflanzen, erreicht jedoch dann den gleichen, konstant bleibenden Wert (in Prozent der Ausgangskonzentration) . Als weiterer Hinweis
Z. PJlanzenphysiol. Bd. 75. S. 277-286. 1975.

282 ]. KNAPPE und D. WERNER
100.--------------------------------, ("10) , ,
'0
. :: \\ ; '" \ \
~~ ~
\ ~~ 0
' ..... - -e----- .-----e-----e-----------
20
o
2 6
A G E
Abb. 3: Abnahme der Jod-Konzentration im Kulturmedium bei Wachstum im Dunkeln von .----. Asparagopsis 0---0 Falkenbergia. Weitere Versuchsbedingungen: s. Abb. 1.
Fig. 3: Decrease of iodine concentration in the culture medium with growth in the dark • - - --. Asparagopsis 0 --0 Falkenbergia. Other data as described in fig. 1.
Tab. 1: Vergleichende Extraktion von 131J-markierten Substanzen aus gefriergetrockneten Thalli von Asparagopsis und Falkenbergia, die vorher mit jeweils gleichen Mengen von 131 J (50 ,uCi 131J/10 ml Nahrlosung) fur 7 Tage inkubiert waren. Die Gesamtaktivitiit in den Thallus-Proben wurde gleich 100 % gesetzt, die angegebenen Prozentwerte bei Asparagopsis und Falkenbergia geben den extrahierten Anteil an.
Extraktionsmittel und Bedingungen
1 n HCI (70 r C, 1 Std .)
aqua bid. (70 °C, 1 Std .)
saures t-Butanol (22 °C, 1 Std.) (t-Butanol : 25 % HCI c= 1000: 3)
saures t-Butanol (60 °c, 1 Std.)
ammoniakal. Methanol (22 c'C, 1 Std.) (Methanol: NH3 = 99 : 1)
Tris-Puffer (30 e, c, 1 Std.) (pH 7,6; 0,05 M)
Z. PJlanzenphysiol. Bd. 75. S. 277-286. 1975.
Asparagopsis
93,66
87,58
15,03
16,32
17,13
36,42
Falkenbergia
53,88
42,89
1,38
2,22
18,46
13,38

Jodstoffwechsel von Asparagopsis und Falkenbergia 283
fiir die lichtunabhangige Aufnahme von 131 J werden Versuche gcwertet, nach denen Photosynthesehemmstoffe wie DCMU (10-6 M) und CCCP (Carbonylcyanid-m-chlorPhenylhydrazon, 5 X 10-6 M) keine signifikanten Unterschiede in der Aufnahme von 131 J aus der Nahrlosung bewirken. Auch bei Laminaria hatte Dunkelheit in einem Kurzzeitexperiment (40 min Dunkelzeit) keinen Einflug auf die Aufnahmegeschwindigkeit (SHAW, 1959).
Die signifikanten Unterschiede in den Aufnahmekinetiken der beiden Generationstypen Asparagopsis-F alkenbergia gab Anlag zur Untersuchung der Frage, ob das aufgenommene Jod in unterschiedlicher Weise in den Thalli gebunden ist. Dies sollte durch vergleichende Extraktionen mit mehreren Losungsmitteln untersucht werden. Die gefriergetrockneten Algen wurden mit den in Tabelle 1 aufgefiihrten Extraktionsmitteln behandelt und der Anteil der Gesamtaktivitat bestimmt, der in die losliche Phase iibergeHihrt werden kann.
Dabei ist aus der Tabelle zu ersehen, dag mit einer Ausnahme die Aktivitat im Oberstand von Asparagopsis-Proben wesentlich hoher liegt als bei den FalkenbergiaProben. Wahrend bei Asparagopsis rund 90 % des aufgenommenen Jods durch eine Extraktion bei 70 C in 1 n HCI oder in a. bidest. moglich ist, werden bei Falkenbergia unter gleichen Bedingungen nur rund 50 % des Jods freigesetzt. Der relative Unterschied wird bei einer Extraktion mit saurem t-Butanol noch wesentlich groger. 1m Vergleich zu Falkenbergia lagt sich hiermit aus Asparagopsis ungefahr achtmal soviel an jodhaltigen Substanzen extrahieren. Nur bei einer Extraktion mit ammoniakalischem Methanol wird aus beiden Thallusformen annahernd gleich viel herausgelost. Die dargestellten Unterschiede lassen sich bereits nach 5miniitiger Extraktionszeit messen. Auch bringt ein Verbleiben des Materials im Extraktionsmittel iiber eine Stunde hinaus nur noch geringfiigige Steigerungen im lOslichen Anteil.
Urn zu sehen, ob durch EiweiBhydrolyse Jod in die losliche Fraktion iiberfiihrt werden kann, wurden die Riickstande einer Pronase- bzw. Trypsinbehandlung unterzogen. Bei Asparagopsis wird durch diese Behandlung rund 20 % der Gesamtaktivitat zusatzlich lOslich, einen etwa gleich groBen Anteil erhalten wir bei F alkenbergia (Abb. 4).
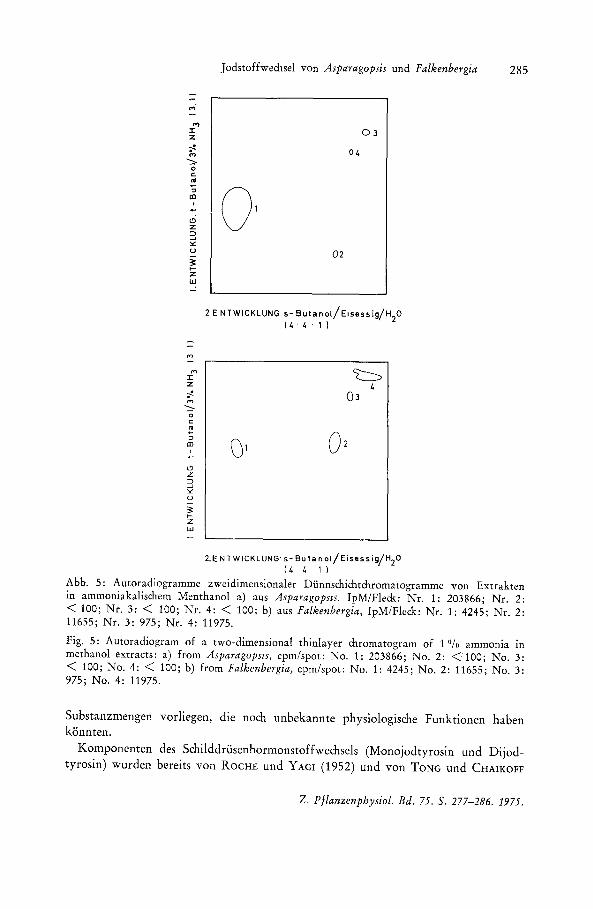
In den bisher dargestellten Experimenten wurden erhebliche Unterschiede bei Asparagopsis und Falkenbergia hinsichtlich der quantitativen Aufnahme von Jodid und der Extraktion jodhaltiger Substanzen nachgewiesen. 1m folgenden untersuchten wir die Frage, ob qualitative Unterschiede in der Zusammensetzung der jodhaltigen Extrakte aus beiden Generationstypen festzustellen sind. In Anlehnung an die fiir Schilddriisenhormone ausgearbeiteten diinnschichtchromatographischen Trennungen wurden die ammoniakalischen Methanolextrakte auf Celluloseplatten getrennt (Abb. 5). Diese Extrakte wurden zunachst gewahlt, weil sie annahernd die gleiche qualitative Extraktion bei beiden Formen ergaben. Wahrend bei dem Extrakt aus Asparagopsis mehr als 99 Ofo der Aktivitat auf einem Fleck des Chromatogramms vorliegt, ist bei F alkenbergia die Markierung deutlich auf 4 Flecken verteilt, wobei die Aktivitat im Verhaltnis 42: 40: 15 : 3 in den einzelnen Flecken gemessen wurde.
Z. Pjlanzenphysiol. Bd. 75. S. 277-286. 1975.

284 ]. KNAPPE und D. WERNER
100,---------------------------------, ('/.)
80
<i
I- 60
>
40
2 4 6 8 9 10 11 12
5 UNO E N
Abb. 4: Pronase- und Trypsin-Behandlung der Sedimente (Fraktionierte Zentrifugation 5 min 1400 X g) eines Gewebeaufschlusses in Tris-Puffer (0,05 M, pH 7,6) von --- Asparagopsis - - Falkenbergia. 6. Inkubation mit Trypsin bci 30 DC, 0 Inkubation mit Pronase bei 30 DC, 0 Kontrolle (Inkubation in Tris-Puffer bei 30 DC). Ordinate: Prozentangabe der nicht sedimentierbaren (1400 X g/5 min) 131 J-Aktivitat bezogen auf die Gesamtaktivitat.
Fig. 4: Treatment with pronase and trypsin of the sediments (centrifugation 5 min at 1400 X g) of tissues, ground in tris-buffer (0.05 M, pH 7.6) --- Asparagopsis -- Falkenbergia. 6. Incubation with trypsin at 30 DC, 0 Incubation with pronase at 30 DC, 0 control incubation in tris-buffer at 30 DC) vertical axis: per cent of 13I I-activity, not sedimented at 1400 >: g for 5 min.
Der sehr kleine, aber autoradiographisch noch eindeutig nachweisbare Fleck. Nr. 3 des Asparagopsis-Extraktes hat im zweidimensionalen Chromatogramm die gleichen RF-Werte wie der Fleck. Nr. 3 des Extraktes aus Falkenbergia, jedoch mit weniger als 0,1 % der relativen Aktivitat. Dagegen sind keine Flecken im Autoradiogramm des Asparagopsis-Extrakts mit den RF-Werten der Flecken Nr. 2 und 4 des Falkenbergia-Extrakts vorhanden, die zusammen ca. 82 % der Gesamtaktivitat ausmachen. Vergleichschromatogramme mit r, Dijod-Tyrosin, Dijod-Thyronin, Trijod-Thyronin, 5-Jod-uridin, 5 Jod-2-desoxyuridin und 5-Jod-uracil zeigten, dag der Fleck. Nr. 4 des Falkenbergia-Extraktes (mit ca. 42 % der Gesamtaktivitat) mit keiner dieser Substanzen identisch ist.
Die Auftrennung wagriger Extrakte (Trispuffer 0,05 M, pH 7,6) aus Asparagopsis und Falkenbergia ergab hinsichtlich der Verteilung der Aktivitat auf einen Fleck bei Asparagopsis (> 99 Ofo) und der Verteilung auf mehrere Flecken bei Falkenbergia ein qualitativ ahnliches Ergebnis wie bei der in Abb. 5 dargestellten Auftrennung. Die chemische Identifizierung der verschiedenen chromatographisch aufgetrennten jodhaltigen Verbindungen erscheint nach den vorliegenden Untersuchungen besonders bei Falkenbergia interessant, da hier mehrere jodhaltige Verbindungen in signifikanten
Z. Pjlanzenphysiol. Bd. 75. S. 277-286. 1975.
..., :I: Z
;! '" -::::,. o c It
:::J III ,
" z ::;) ...J >< U
~ .... z UJ
--'" -'" :I:
Z
;! '" ~ 0 c .. " III
" z 3 "" u
~ .... z UJ
Jodstoffwechsel von Asparagopsis und Falkenbergia 285
03
04
0, 02
2 E NTWICKLUNG 5- BUlanOI/ElseSS ig/H20
( 4 . 4 . I )
~ 4
03
C} 02
2.E N T W ICKLUNG' 5- Bu la n 01/ Eis.55 ig/H20 (4 4 I)
Abb. 5: Autoradiogramme zweidimensionaler Diinnschichtchromatogramme von Extrakten in ammoniakalischem Menthanol a) aus AsparagopslS. IpM/Fleck : Nr. 1: 203866; Nr. 2: < 100; Nr. 3: < 100; Nr. 4: < 100; b) aus Falkenbergia, IpM/Fleck: Nr. 1: 4245; Nr. 2: 11655; Nr. 3: 975; Nr. 4: 11975.
Fig. 5: Autoradiogram of a two-dimensional thinlayer chromatogram of 1 % ammonia in methanol extracts: a) from Asparagopsls, cpm/spot: No.1: 203866; No.2 : <100; No.3: < 100; No.4: < 100; b) from Falkenbergia, cpm/spot: No.1: 4245; No.2: 11655; No.3: 975; No.4 : 11975.
Substanzmengen vorliegen, die noch unbekannte physiologische Funktionen haben konnten.
Komponenten des Schilddriisenhormonstoffwechsels (Monojodtyrosin und Dijodtyrosin) wurden bereits von ROCHE und Y AGI (1952) und von TONG und CHAIKOFF
z. Pflanzenphysiol. Bd. 75. S. 277-286. 1975.

286 ]. KNAPPE und D. WERNER
(1955) bei Braunalgen nachgewiesen. Eine vollstandige quantitative Aufklarung des Jodstoffwechsels bei einer Rotalgen- oder Braunalgenart ist jedoch bis heute nicht erfolgt. Diese Aufklarung ware die Voraussetzung, urn die physiologischen Funktionen jodhaltiger Metabolite bei Algen im Zusammenhang mit dem Generationswechsel zu klaren.
Literatur
BAILY, N. A., and S. KELLY: Iodine exchange in Ascophyllum. Bio!. Bul!. 109, 13-21 (1955). FELDMANN, ]. und G.: Recherches sur les Bonnemaisoniacees et leur alternance de genera
tions. Ann. Sc. nat. Bot. 3, 75-175 (1942). FRIES, L.: Influence of iodine and bromine on growth on some red algae in axenic culture.
Physiol. Plant. 19, 800-808 (1966). HSIAO, S. 1. c.: Life history and iodine nutrition of the marine brown alga Petalonia fascia.
Can. ]. Bot. 47, 1611-1616 (1969). KELLY, S., and N. A. BAILY: The uptake of radioactive iodine by Ascophyllum. BioI. Bull.
100, 188-190 (1951). KLEMPERER, H. G.: The accumulation of iodide by Fucus ceranoides. Biochem. ]. 67, 381-390
(1957). KYLIN, H.: Ober die Blasenzellen einiger Florideen und ihre Beziehung zur Abspaltung von
Jod. Ark. f. Bot. 14, 1-13 (1915). ROCHE, ]., et Y. YAGI: Sur la fixation de l'iode radioactif par les algues et sur les constituants
iodes des Laminaires. Compt. rend. Soc. bio!. 146, 642-645 (1952). SHAW, T. I.: The mechanism of iodide accumulation by the brown sea weed Laminaria
digitata. Proc. Roy. Soc. London, Ser. B 150, 356-371 (1959). V. STOSCH, H. A.: Wirkungen von Jod und Arsenit auf Meeresalgen in Kultur. Proc. 4th
Internat. Seaweed Symp., 142-150 (1963). TONG, W., and I. L. CI-lAIKOFF: Metabolism of F3! by the marine alga, Nereocystis luetkeana.
]. bio!. chern. 215, 473-484 (1955). WOOLERY, M. L., and R. A. LEWIN: Influence of iodine on growth and development of
the brown alga Ectocarpus siliculosus in axenic culture. Phycologia 12, 131-138 (1973).
Dr. JOHANNA KNAPPE, Prof. Dr. D. WERNER, Fachbereich Biologie, Botanisches Institut der Universitiit, D-355 Marburg/Lahn, Auf den Lahnbergen.
Z. Pflanzenphysiol. Bd. 75. S. 277-286. 1975.





























