Embed Size (px)
Citation preview

JOURNAL FtJR
O R N I T H O L O G I E
B a n d 108 1967 N r . 3
Aus dem Zoologischen Institnt der Humboldt-Universit~t Berlin
Probleme der Motivwahl und der Gesangsaktivitiit bei PhyUoscopus trochilus ( L . )
Vou Midlael Schubert, Berlin
A. Einleitung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 265
B. Methoden und Material . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 267
C. Der EinfluB des nachbarlichen Gesanges auf Motive und Gesangsaktivit~it . . . 271
D. Die Tageszeit als regelnder Faktor . . . . . . . . . . . . . . . . . . . . . . . 273
1. Gesangsbeginn und -ende . . . . . . . . . . . . . . . . . . . . . . . . . . 273
2. Die Gesangsaktivit~it im Tageslauf . . . . . . . . . . . . . . . . . . . . . 273
3. Tageszeitliche Unterschiede in der HKufigkeit yon Variationen . . . . . . . 274
4. Variationen in der Strophenl~nge . . . . . . . . . . . . . . . . . . . . . . 277
E. Modifikationen wiihrend des Revierkampfes . . . . . . . . . . . . . . . . . . 277
F. Der brutzeitliche Status als rege]nder Faktor . . . . . . . . . . . . . . . . . . 278
1. Die Beeinflussung der GesangsaktivitKt . . . . . . . . . . . . . . . . . . . 278
2. Unterschiede in tier HSufigkeit yon Variationen . . . . . . . . . . . . . . 279
3. Die H/iufigkeit yon ,,Sirren" und ,,sit"-Lauten . . . . . . . . . . . . . . . 281
4. Variationen in der Strophenl/inge . . . . . . . . . . . . . . . . . . . . . . 282
G. Einflug der Singwarte . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 283
H.. Diskussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 284
I. Zusammenfassung, Summary . . . . . . . . . . . . . . . . . . . . . . . . . . 288
J. Literaturverzeichnis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 290
A. Einleitung In den letzten Jahrzehnten behandel ten zahlreiche Autoren die Gesangsaktivi-
t/~t bei verschiedenen Passeres ira Hinblick auf exogene und endogene Bezugs-
systeme. Besonders wurde die Steuerung yon Beginn und Ende des Vogelgesan-
ges und seine Intensit/it (Aktivit/it) im Tagesverlauf beachtet. Schon HAECKER
(1900, 1916, 1924) und sein Schiller SCHWAN (1920, 1921--23) untersuchten die
, ,Reizphysiologie" des Vogelgesanges im Hinblick auf Tageszeit und meteorologi-
sche Faktoren. Ferner sei hingewiesen auf WRIGHT (1913), ZIMMER (1920), PAL~-
GaEN (1932) und seine Schule, DORNO (1924), ALLARD (1930, 1934), KUUSISTO
[ J. Orn. 2 6 6 MICHAEL SCHUBERT [ 108
(1941), SCHEE~ (1941), NICE (1943), FRANZ (1943, 1949), GROEBEELS (1956), WAGNER (1956), MEa~EL (1956, 1958), CvaIO (1959), HO~MANN (1959), LEOPOLD
E~NON (1961), VnI~M (1961). ASCHOF~ und WEVEa priizisierten die Befunde yon Beginn und Ende der Tagesaktivit~t beiVSgeln unter Zuhilfenahme eines kyberne- tischen Modells (AscHo~ a WEVEn 1962). Die jahreszeitlichen Aktivit~itsschwankun- gen werden in den hShcren geographischen Breiten auf dem Wege fiber gonadotrope Hormone im wesentlichen yon der Tagesliinge gesteuert [BSKER (1923), F. A. H. MARSHALL (1936), BURGER (1940, 1947, 1949, 1953), WOL~SON (1941, 1952), SAUNDERS (1947, 1948 a und b), FRANZ (1949), ST~EVE (1950), A.J. MARSHALL (1951, 1952, 1960), DAVIS & DAVIS (1954), LETHONEN (1954), CUNNINGHAM (1955), FAa~En (1959) U. m].
Methodisch ungleich schwerer und aufwendiger ist die Untersuchung jahres- und tageszeitlicher q u a 1 i t a t i v e r Ver~inderungen in Ausdruek und Motiven yon Vogelges~ngen durchffihrbar. Dank der heute sear pr~zise arbeitenden spek- trographischen Analyseverfahren bietet der Gesang als se,~r rein strukturiertes Ausdrucksverhalten giinstige Ansatzpunkte, um die Zusammenhiinge zwischen spezifischem Aktivit~tsspiegel und dem Wechsel in der Struktur des Signals zu untersuchen. Jahres- und tageszeitliche _~nderungen der Gesangsqualiti~t sind bis- lang seAr vernachUissigt oder nur als beiliiufiges Untersuchungsergebnis behan- delt worden (Literatur s. Diskussion).
: i . . . . . .
:~^~-~ ~ i ? : - ~ : ~ , , - ~ .............. j :.-: ........ ~, • ,~ ~:~ ~:.... " .:: :,;~ ..... n>:,~'~,:.:.::',~7,' .,~:;~-~ .'.~:~.~'~: "~ ..... = ~,- ....... ~.
"~I'\, ",, ",, " ,~,~%~i .~i~,"., " ,." ,," ,, ",," ,, "I'~~.~ | .... '~'I,, ~.'c~ .... /~:~!.~-!'~.~;~ i~ ........ " ..... I I ~ ~ - : ~
- ~ , , ~ ' ~ ~ - . .... - - ~ ~ i .~,~,,!-~ ~: .;; ,, ,, ,, :::...
~ ' ,',':.: .~:-: ..,,. ~':~,~ ~~;~::.!: A: " ~'."'5 : .... ~N;;~-, ~;!/~ ~ ;X ".".. ".,",, ",,",,"., ," ;i;>:.~:!;
• WegGraben ..... ,,, ,,, .~,:..,_~. ............ '., ",, :=L:~~::~
" o. ~ ~,~ ~ ' . ' . ,." .." ,,;.;:,;~:?~:~"
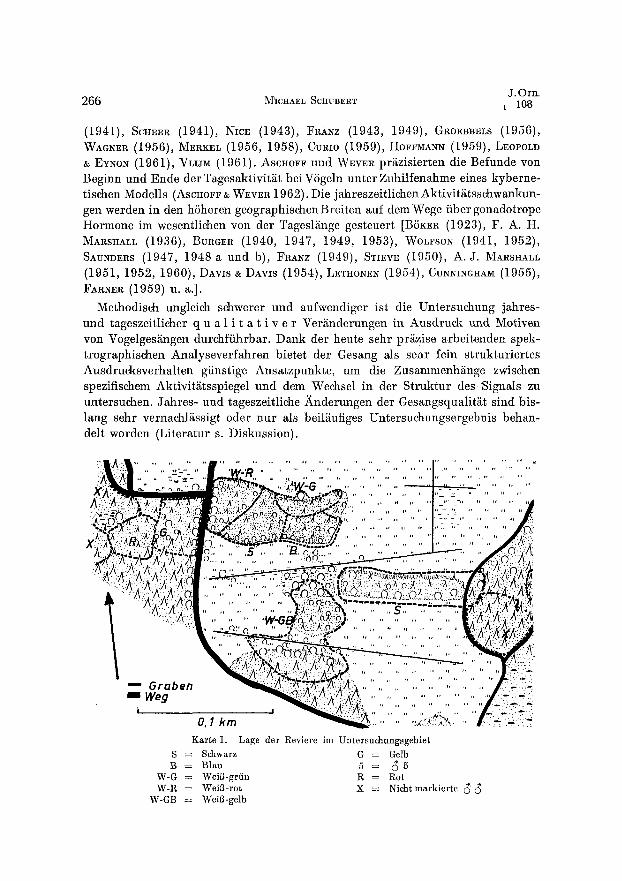
Karte 1. Lage der Reviere im Untersuchungsgebiet
S = Schwarz G = Ge/b B = Blau 5 = ~ 5
W-G = WeiS-griin R ---- Rot W-R = Weil]-rot X : N ich tmark i e r t e ~ (~
W-GB = Weil]-gelb

Heft 3] 1967 J Gesang des Fitis 267
Als Untersuchungsobjekt fiel die Wahl auf Phylloscopus trochilus, der in der
Mark Brandenburg ein hiiufiger Brutvogel ist. Sein Gesang ist s t rophig und reich an Themen, und auL3erdem lieflen die nahe Verwandtsehaft zu dem sympatr isehen
Ph. collybita und die hiervon bestimrnte Auspr / igung des Gesanges als ar t isol ie- rende S igna l s t ruk tur sowie die vorrangige Bedeutung der akustisehen Kommuni-
kat ionsmit te l im Leben dieses Vogels den Fi t i s besonders geeignet erseheinen. Eine ausfiihrliehe Besehreibung des Gesanges anhand von Sonagrammen l ieferten
THIELCKE & LINSENMAIR (1963). Neben der Aufzueht von K a s p a r Hausern 3. Ord- nung und der KEfighaltung adul ter Mgnnchen wurde vor allem der Motivgesang
im Revier yon Anfang Apr i l bis zur Sommermauser untersucht.
B. M e t h o d e n u n d M a t e r i a l
Eme kleine Population yon Fitislaubs~ingern nSrdlieh yon Bernau bei Berlin (52 °', 45" N, 13 °, ~6' E) wurde von Anfang April bis Anfang Juli 1'964 systematiseh untersucht. Die Reviere lagen grol]ent~ils in l ljiihrigen Kiefernsehonungen mit eingesprengten Birken (Betula pubescens Ehrh.), Stieleiehen (Quercus robur L.) Robinien (Robinia pseud- acacia L.) und Pfaffenhfiteben (Evonymus europaea L.). Infolge der inselartigen Aus- dehnung der Anpflanzungen waren die einzelnen Territorien der insgesamt 12 unte~- suehten M/innehen dureh Weideland und Wege meist deutlieh getrennt. (Karte 1). Von diesen Exemplaren wurden bevorzugt beobaehtet und auf Band aufgenommen:
1. d 5 2. Sehwarz, 3. Gelb, 4. Blau, 5. WeiB-gelb, 6. Rot, 7.
Revier besetzt seit 13. IV., Brutvogel Revier besetzt seit 15. IV., blieb unverpaart Revier besetzt seit 17. IV., Brutvogel Revier besetzt seit 20. IV., Brutvogel Revier besetzt seit 20. IV., Brut im Juni Revier besetzt seit 24. IV., Brutvogel
WeiB-griin, Revier besetzt seit 29. IV., blieb unverpaart
Die d Sehwarz, Weil3-gelb und Blau wurden in regelmiigigen Abstiinden an mehreren Tagen auf Tonband aufgenommen: morgens allstiindlieh bis 8 Uhr, mittags zwisehen 12 und 14 Uhr und abends meist ab 17 Uhr. Hinzu kommen zahlreiehe Aufnahmen zu anderen Zeiten und yon anderen Hiihnen. Ab 7. Mai wurden naeh demselben Turnus 15-minfitige Ziihlungen der dem GehSr naeh unterseheidbaren Gesangsvariationen und der Ges/inge iiberhaupt (n/15 rain.) durehgefiihrt. Die Resultate wurden Ms Graphik dem tiigliehen Beoba&tungsprotokoll zur weiteren Auswertung beigeffigt. Diese enth~ilt ferner meteorologisehe Daten (Temperatur, rel. Luftfeuehte, Windst~irke und -riehtung, BewS1- kungsgrad, Sonnenseheinautogramm) sowie Messungen der Liehtintensitiit unmittelbar vor jeder Tonbandaufnahme und jedem 15-Min.-Test.
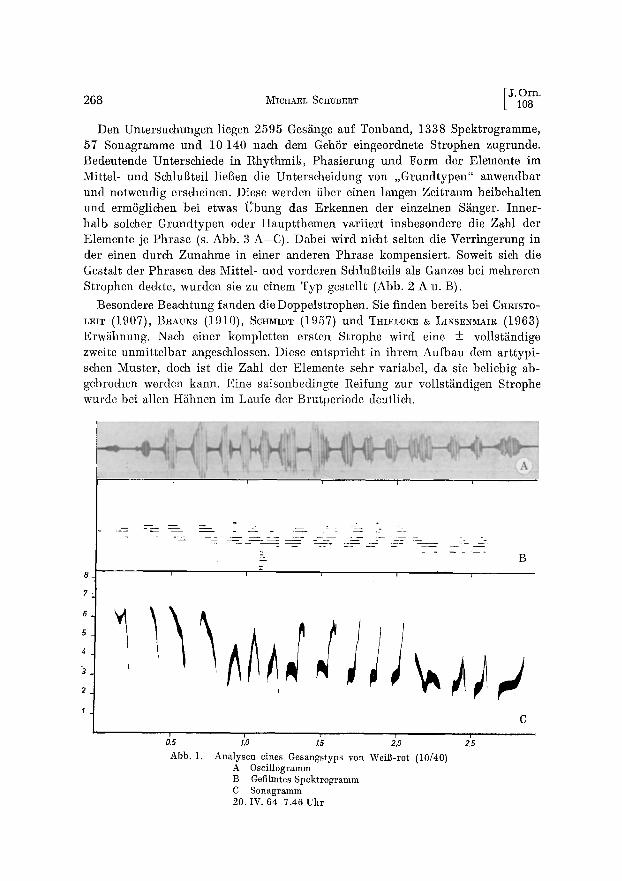
Zur Aufnahme verwendete ieh das Transistorger~it Gau~Dm TK 4 mit dem Mikrofon DM 121 und einem paraboloiden Reflektor. Die Analyse erfolgte mit einem Tonfrequenz- spektrometer (SSP-10, Funkwerk KSpeniek, 40-17250 Hz) und einem Filmreeorder bei halber Bandgesehwindigkeit (Besehreibung des Verfahrens bei TEr~BROCK 1963). Die Methode erwies sieh als brauchbar fiir den Vergleieh der Strophen untereinander, war abet ungeeignet ffir Angaben fiber die Sehallstiirke und eine genaue Frequenzbestimmung (Abb. 1 B). Einige der priignantesten Beispiele wurden sehliel]lieh noeh sonagraphiert. Die Gesangsliinge wurde mit der Stoppuhr bei halber Bandgesehwindigkeit gemessen.
[J.Orn. 268 ~V[ICHAEL SCHUBERT L 108
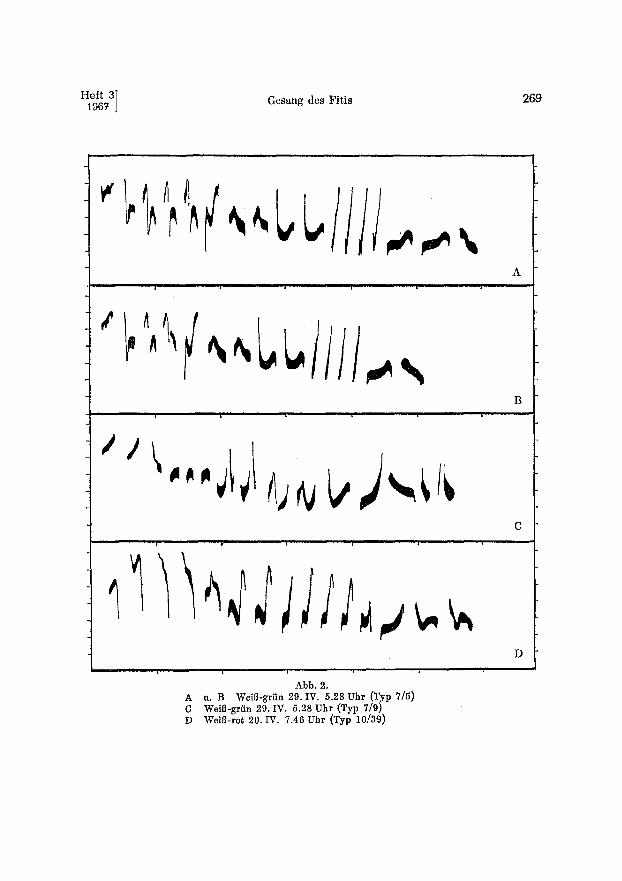
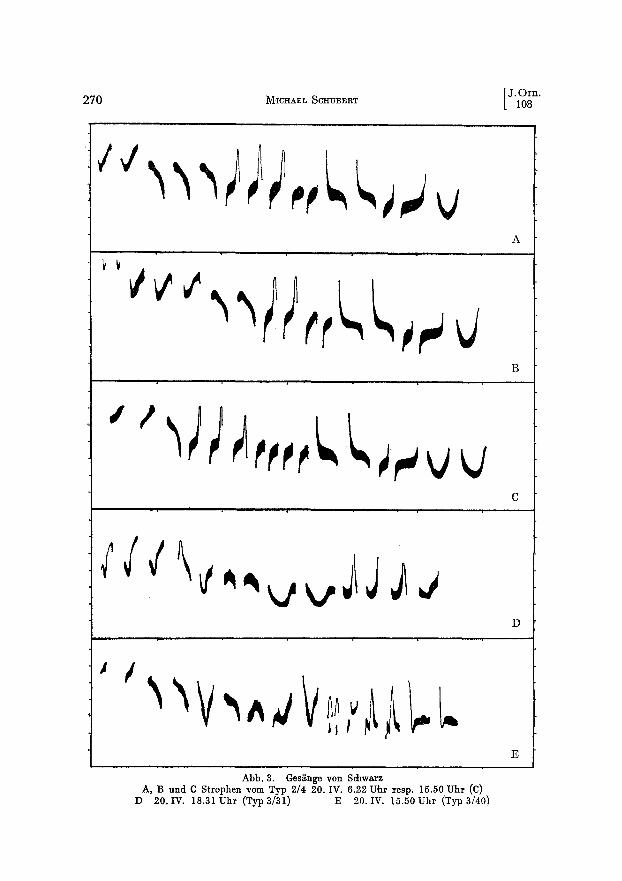
Den Untersuchungen liegen 2595 GesBnge auf Tonband, 1338 Spektrogramme, 57 Sonagramme und 10 140 nach dem GehSr eingeordnete Strophen zugrunde. Bedeutende Unterschiede in Rhythmik, Phasierung und Form der Elemente im Mittel- und SchluBteil lieBen die Unterseheidung yon ,,Grundtypen" anwendbar und notwendig erscheinen. Diese werden fiber einen langen Zeitraum beibehalten und ermSglichen bei etwas CTbung das Erkennen der einzelnen Siinger. Inner- halb soleher Grundtypen oder Hauptthemen variiert insbesondere die Zahl der Elemente je Phrase (s. Abb. 3 A--C). Dabei wird nicht selten die Verringerung in der einen dutch Zunahme in einer anderen Phrase kompensiert. Soweit sich die Gestalt der Phrasen des Mittel- und vorderen SchluBteils als Ganzes bei mehreren Strophen deekte, wurden sie zu einem Typ gestellt (Abb. 2 A u. B).
Besondere Beachtung fanden die Doppelstrophen. Sie finden bereits bei CHRISTO- L~IT (1907), BnAVNS (1910), SCHMIDT (1957) und THIELCKE & LINSENMAIR (1963) Erw~hnung. Nach einer kompletten ersten Strophe wird eine +- vollst~indige zweite unmittelbar angeschlossen. Diese entspricht in ihrem Aufb~u dem arttypi- sehen Muster, doch ist die Zahl der Elemente sehr variabel, d~ sie beliebig ab- gebroehen werden kann. Eine saisonbedingte Reifung zur vollstiindigen Strophe wurde bei allen H~hnen im Laufe der Brutperiode deutlich.
- -£_
8
7
6
5
2
1
I I I I
~,s ;o 15 ~,o 215 Abb. 1. Analysen eines Gesangstyps yon Wei]-rot (10/40)
A Oscillogramm B Gefilmtes Spektrogramm C Sonagramm 20. IV. 64 7.46 Uhr
C
Heft 3] 1967 J
Gesang des Fitis 269
i I L I I I I
Abb. 2. A u. B WeiB-griin 29. IV. 5.28 Vhr !Typ 7/5) C WeiB-gr/in 29. IV. 5.28 Uhr (Typ 7/9) D WeiB-rot 20. IV. 7.46 Uhr (Typ 10[39)
A i
B
C
D
270 MICHAEL SCHUBERT J.Orn.
108
[
A
B
D
E
Abb. 3. Ges~nge yon Schwarz A, B und C Strophen vom Typ 2/4 20. IV. 6.22 U,hr resp. 15.50 Uhr (C)
D 20. IV. 18.31 Uhr (Typ 3/31) E 20. IV. 15.50 Uhr (Typ 3/40)

Heft 3] 1967 J Gesang des Fitis 271
Die Themen beider Teile des Liedes variieren unabh~ngig voneinander. Die yon THIELCKE ~5 LINSENMAIR (1963) untersuchten zweiten Strophen sollen mit hSheren Fre- quenzen als in der ersten Strophe beginnen. Der SchluBteil der 1. Strophe leitet nicht selten mit ansteigenden Schlul]lauten zu dem hohem Anfang der zweiten fiber; dennoeh besteht ein beachtlicher Frequenzunterschied.
Aus Beobachtungen und Aufnahmen wurde ersichtlich, dab die gesamte Va- riabilit~t gewissen Sehwankungen unterliegt. Als MaB fiir die Variationsbreite wurde die H~ufigkeit yon Wiederholungen in Prozenten aller Ges~/nge einer Auf- nahme (meistens 15--20) gewertet. Sie wurde WiederholungsgrSBe oder W°/0 ge- nannt. Je einfSrmiger ein Vogel singt, um so grSBer ist W°/o.
W°/o = (ntl--1) + (nt2--1) . . . . . ÷ (nti--1) • 100
]]at • a
a = Anzahl der Ges~nge der betreffenden Aufnahme
n t l . . i = Anzahl der Ges~iage vom Typ t l . . i
mt = Anzahl untersehiedlicher Typen der Aufnahme
Herrn Prof. Dr. TEMBROCK, ~neinem verehrten Lehrer, verdanke ich das Thema meiner Diplomarbeit, die hier in Auszfigen zugrunde liegt. Prof. TSMBmJCK stellte mir zu Ver- gleichszwecken eigene Bandaufnahrnen, Literatur und die technischen Ger~tschaften des bioakustischen Labors des Zoologischen Institutes zur freien Verffigung. Herr Dipl. Biol. LIPPERW stand mir zeitweilig bei der Gel~indearbeit zur Seite. Die Mitarbeiter am Phone- tischen Institut der Humboldt-Universit~it (Direktor Prof. Dr MEIEr) berieten und halfen in manchen teehnischen Fragen, Herr KYRITZ half bei der Herstellung der Sonagramme, Herr MErZ~ kopierte einen Tell der Tonb~nder. Die Wetterdienststelle Potsdam lieh die meteorologischen MeI3ger~ite aus. Herr Dr. G TmESCK~, Radolfzell, stellte mir Bandauf- nahmen schwedischer und westdeutscher Fitisse und spanischer Weidenlaubs[inger zur Verftigung.
Ohne die groBe Hilfe und Riicksichtnahmo meiner Eltern w~ire die Durchffihrung der Arbeit undenkbar gewesen. Ihnen allen gilt mein herzlicher Dank.
C. D e r Einflufl d e s nachbar l i chen G e s a n g e s auf M o t i v e u n d G e s a n g s a k t i v i t ~ i t
Auf Karte 1 ist die endgiiltige Besiedlung des Beobachtungsgebietes (Anfang Mai 1964) dargestellt. Die Singwarten yon Schwarz, WeiB-gelb, Blau und M. 5 z. B. lagen bei Windstille im gegenseitigen HSrbereich. Da benachbarte Fitislaub- s~nger bekanntlich sehr h~ufig genau alternierend singen, war zu vermuten, dab sich Nachbarn gegenseitig in ihren Variationen beeinflussen kSnnen. Augleichung h~tte eine Verminderung des individualspezifischen Charakters der Strophe zur Folge, Differenzierung w/irde das Erkennen des Nachbarn erleichtern und w~re informationstheoretisch zu fordern. Benachbarte Individuen singen meist grund- verschieden. Es konnte kein Gesangstyp gefunden werden, der "vSllig identisch mit einem Motiv des Nachbarn war. Ahnlichkeiten untereinander stellen sich offenbar ~iul3erst selten gleichzeitig ein, sondern sie treten, falls iiberhaupt, auf, wenn das Motiv des Nachbarn ,,ungebr~uchlich" geworden ist, und zwar einige Tage oder Wochen sp~ter.
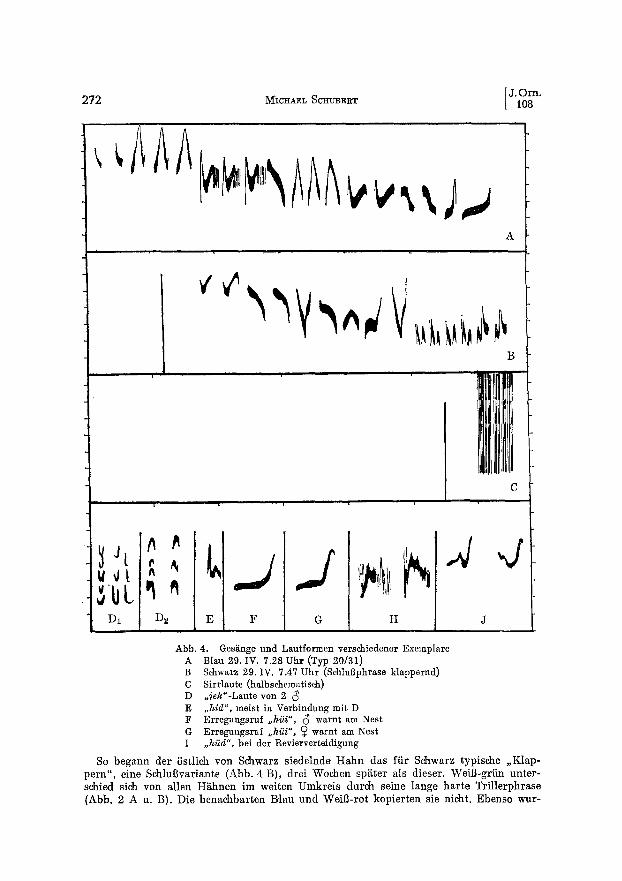
272 ~ICHAEL SCHUBERT
A p
B
C
Abb. 4. Gesgnge und Lautformen verschiedener Exemplare A Blau 29. IV. 7.28 Uhr (Typ 20131) B Schwarz 29. IV. 7.47 Uhr (Sehlu0phrase klappernd) C Sirrlaute (halbschematis,eh) D ,,~ek"-Laute yon 2 (~ E ,,hid", meist in Verbindung mit D F Erregungsruf ,,hi~i", (~ warnt am Nest G Erregungsruf ,,hi~i", ~ warnt am Nest I ,,hiid", bei der Revierverteidigung
So begann der 5stlich yon Schwarz siedelnde H a h n das fiir Schwarz typische ,,Klap- pern" , eine SchluBvariante (Abb. 4 B), drei Wochen sp~ter als dieser. WeiB-griin unter- schied sich yon allen H~ihnen im weitcn Umkreis durch ssine lange har te Tri l lerphrase (Abb. 2 A u. B). Die benachbar ten Blau und W~i~-rot kopierten sie nicht. Ebenso wur-

Hef~ 73 ] des Fitis 273 Gesmug
den der besonders auff~lli~e Typ 20/31 (Abb. 4 A) und andere Themen von Blau weder yon Weil3-griin noch von (~ 5 aufgegriffen.
~hnlichkeiten von Variationen ~iul]ern sich meistens in den Schlul3elementen. In allen Fallen konnte aber kein eindeutiger Hinweis auf echte Anpassungserschei- nungen, die durch die Nachbarschaft bedingt sind, festgestellt werden. So han- delte es sich entweder um Exemplare, die aul3erhalb ihres HSrbereiches siedelten odor deren StrophenschluI3 sporadisch im Umkreis yon mehreren Kilometern zu hSren war.
ROWAN (1955) und THORPE (1958) stellten dagegen lest, dab ein Vogel, der seinen Rivalen ansingt, dazu neigt, mit dem Motiv zu antworten, das dem des Nachbarn am ~hnlichsten ist. Naeh SAUSR (1955 a) gleichen sich die Motive sich ansingender Dorngrasmiicken (Sylvia communis) nach einiger Zeit an (s. auda LANYON, 1957, fiir Sturnella magna).
Das alternierende Singen zweier H~hne hat zwangsl~ufig zur Folge, dab beide das Niveau ihrer Gesangsaktivit~it weitgehend anpassen. Hierbei kann besonders nach der Reviergrfindung Stimmungsfibertragung mitspielen, w~hrend die anf~ng- liche Rivalit~it allm~hlich abflaut.
Beim Ansingen ist die Gesangsaktivit~t, ebenso wie nach Grenzk~mpfen, beson- ders hoch. MAY (1949) fiihrt aus, dab Grenzverletzer sich in der Regel still und unauff~llig im fremden Gebiet bewegen. Diese Feststellung kann ich nicht be- sti~tigen. Wenn CONDER (1948) feststellt, dab die singfreudigsten Stieglitze (Car- duelis carduelis) die grSl]ten Reviere besitzen, so findet man diese Verh~iltnisse bei Phylloscopus trochilus nicht vor.
D. Die Tageszeit als regelnder Fakto~
1. Gesangsbeginn und -ende
Der Gesangsbeginn des Fitislaubs~ngers streut zeitlich nnd in Abh~ingigkeit yon Helligkeitswertcn weniger als das Gesangsende: Minimum 3 Lux (10.6.), Minimum 110 Lux (16.7.). Die grSBte zeitliche Abweichung vom Sonnenaufgang wurde mit - -42 rain. und die geringste mit -+ 0 rain. gemessen. Durchschnitts- werte ffir April und Mai sind 25 Lux und -- 26,4 min. Die zeitlichen Messungen weisen eine geringere Streunng als die Lichtwerte auf.
Die grSBte gemessene Lichtstiirke fiir den letzten Gesang betr~igt 1520 Lux (4. 8.), die niedrigste 37 Lux (24.4.). Die grSBte negative Abweichung vom Zeit- punkt des Sonnenunterganges war - -14 rain. (4.8.), w~hrend die maximale (ne- gative) Abweichung + 20 rain. (2.5.) betr~gt. Der Durchschnittswert fiir April und Mai liegt bei 72,3 Lux und -- 13,75 rain.
2. Die Gesangsaktivit~it im Tageslauf
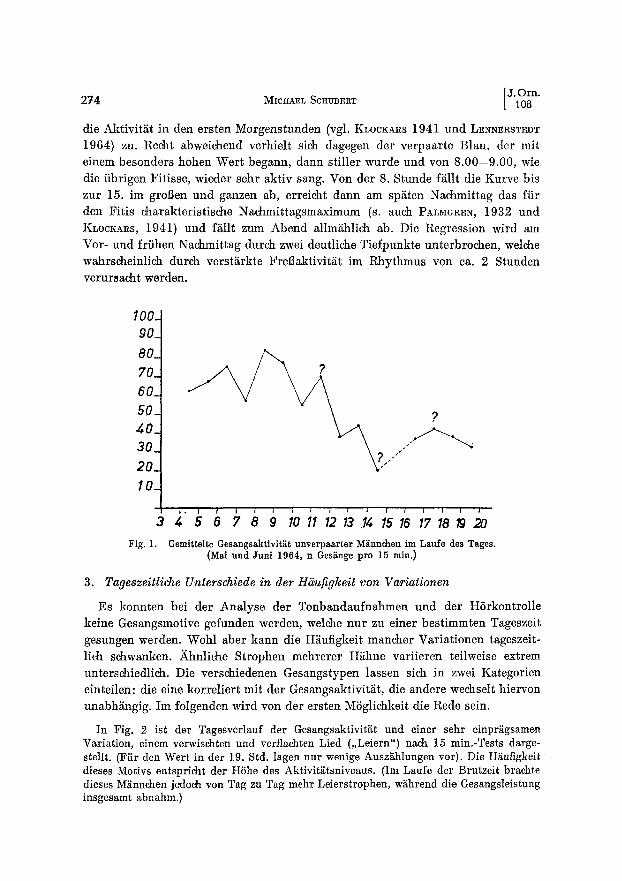
Fig. 1 zeigt die Gesangsaktivitiit im Tagesgang bei unverpaarten M~nnchen. Als Mal3einheit wurde die Anzahl der Ges~nge je 15 min. gew~hlt. In der Regel nimmt
[J.Orn. 274 MIem~ET SCHUBERT [ 1.08
die Aktivitiit in den ersten Morgenstunden (vgl. KLOCKARS 1941 und LENNERSTEDT 1964) ZU. Recht abweichend verhielt sieh dagegen der verpaarte Blau, der mit einem besonders hohen Wert begann, dann stiller wurde und von 8.00--9.00, wie die iibrigen Fitisse, wieder sehr aktiv sang. Von der 8. Stunde fiillt die Kurve bis zur 15. im groBen und ganzen ab, erreicht dann am sp~iten Nachmittag das fiir den Fitis charakteristische Nachmittagsmaximum (s. auch PALMGREN, 1932 und KLOCKARS, 1941) und fiillt zum Abend allmiihlich ab. Die Regression wird am Vor- und friihen Nachmittag durch zwei deutliche Tiefpunkte unterbrochen, welehe wahrseheinlich durch versti~rkte Frel]aktivit~t im Rhythmus von ca. 2 Stunden verursacht werden.
100. 90
80
70 60
50 40 30
20
10_
?
?
9io' ' ' ' i s ' . . . . 3 4 11 12 13 14 16 17 18 19 20
Fig. I. Gemittelte Gesangsaktivit~t unverpaarter M~nndlen im Laufe des Tages. (Mal und Juni 1964, n Ges~nge pro 15 rain.)
3. Tageszeitliche Unterschiede in der Hiiufigkeit von Variationen
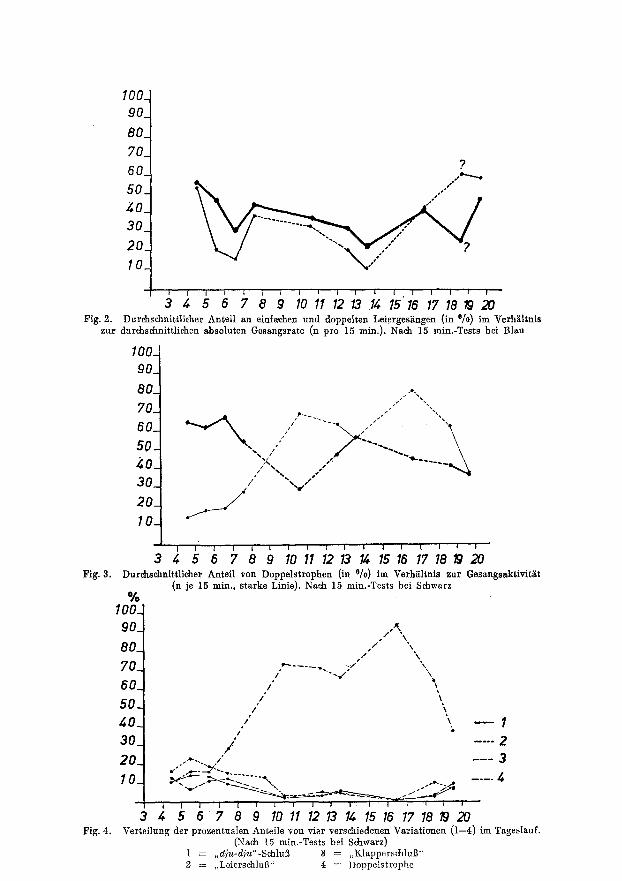
Es konnten bei der Analyse der Tonbandaufnahmen und der HSrkontrolle keine Gesangsmotive gefunden werden, welche nur zu einer bestimmten Tageszeit gesungen werden. Wohl aber kann die H~iufigkeit maneher Variationen tageszeit- lich schwanken. Ahnliche Strophen mehrerer Hiihne variieren teilweise extrem untersehiedlieh. Die verschiedenen Gesangstypen lassen sich in zwei Kategorien einteilen: die eine korreliert mit der Gesangsaktivit~t, die andere wechselt hiervon unabhiingig. Im folgenden wird yon der ersten MSglichkeit die Rede sein.
In Fig. 2 ist der Tagesverlauf der Gesangsaktivit~t und einer sehr einpr~igsamen Variation, einem verwischten und verflachten Lied (,,Leiern") nach 15 min.-Tests darge- stellt. (Ffir den Wert in der 19. Std. lagen nur wenige AuszSJalungen vor). Die H/iufigkeit dieses Motivs entspricht der HShe des Aktivit~tsniveans. (Im Laufe der Brutzeit brachte dieses Miinnchen jedoch von Tag zu Tag mehr Leierstrophen, w~ihrend die Gesangsleistung insgesamt abnahm.)
100 90
80 70 60 50 Z,O_ 30_
20 10
? • sess~ ~ S $ ) ~
~ D J ~ ~ ~ 9 ~ , )2 ) J' ] 5 . J ~" ~ , ) - ,4 5 8 0 11 13 1̀ 4 16 17 18 19 20
Fig. 2. Durchschnittlicher Anteil an einf~chen und doppelten Leierges~ngen (in °/o) im Verh~ltnis zur durchschnittlichen absoluten Gesangsrate (n pro 15 rain.). Nach 15 min.-Tests bei Blau
100_ 90
80 ... 7 0 , - . / "...,
SO .. / /
I ~ js 30_ ..,
20_ _ . ~ 10_ ~
I ; ' ' ' ' 3 `4 5 10 11 12 .1̀ 4 15 17 18 19 20 Fig. 3. Durchschnittlidler Anteil yon Doppelstrophen (in °/o) im Verh~iltnis zur Gesangsaktivit~it
(n je 15 rain., starke Linie). Nach 15 rain.-Tests bei Schwarz %
100_ 90_
80_ 70 60 50 `40_ 30_
20_
10_
Fig. 4.
/ • \
/ s , \
/ \ / \
/ \ / " \
/ 1
. . . .
~ m 3
. . . . `4
3 ,4 5 6 7 8 9 1011 1213 lZ, 1516 17181920 Verteilung der prozentualen Anteile yon vier verschiedenen Variationen (1--4) im Tageslauf.
(N~ch 15 min.-Tests bei Schwarz) 1 = ,,d}u-dju"-Schlu] ] = ,,Klappers,chluB" 2 = ,,Leiersch]u•" 4 = Doppelstropbe
276 MICHAEL SCHUBERT
% 100_ 90_
80_ 70_ 60_ 50_ ,~0
10_
5. 15. 2'5. ,~. 1,~. 2~.
Mai Jun i
Fig. 5. Prozentuale Hgufigkeit verschiedener Variationen yon Schwarz (s. Fig. 4) und die jeweilige Gesangsaktivit~t (n pro 15 min.). Nach 15 min.-Tests 4.00 bis 7.00 Uhr im Mai und Juni
Fig. 6.
w(%) 60_ 50_ ,~0_ 30
20_ 10_
11 13 .15 15 16 17 18 19 20
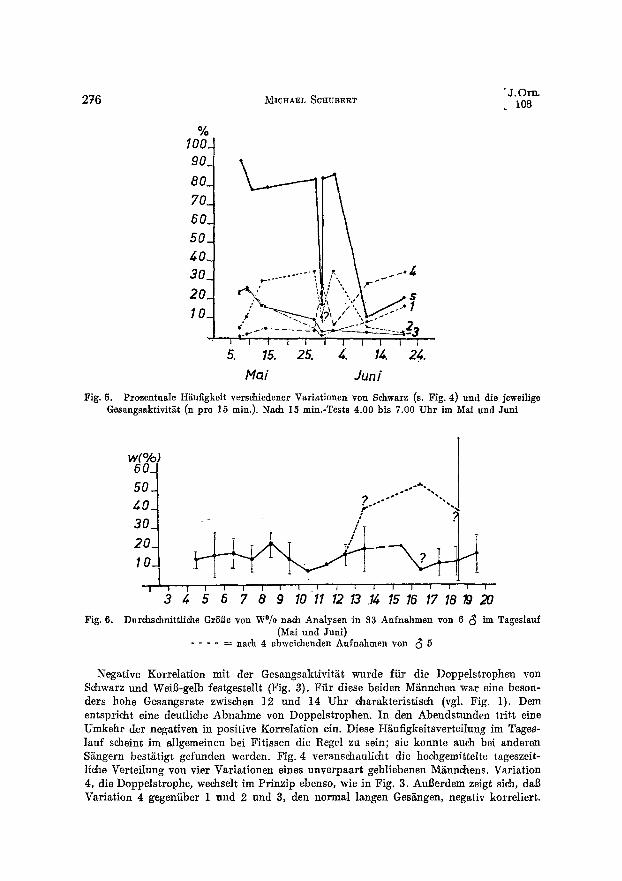
Durchschnittliche GrSl3e yon W°/o nach Analysen in 83 Aufnahmen yon 6 (~ im Tageslauf (Mai und Juni)
. . . . . nach 4 abweichenden Aufnahmen yon (~ 5
Negative Korrelation mit der Gesangsaktivit/it wurde ffir die Doppelstrophen von Schwarz und WeiB-gelb festgestellt (Fig. 3). Fiir diese beiden MEnnchen war eine beson- ders hohe Gesangsrate zwischen 12 und 14 Uhr eharakteristisch (vgl. Fig. 1). Dem entspricht eine deutliche Abnahme yon Doppelstrophen. In den Abendstunden tritt eine Umkehr der negativen in positive Korrelation ein. Diese H£ufigkeitsverteilung im Tages- lauf scheint im allgemeinen bei Fitissen die Regel zu sein; sie konnte auch bei anderen SEngern best~tigt gefunden werden. Fig. 4 veranschaulicht die hoehgemittelte tageszeit- liche Verteilung yon vier Variationen eines unverpaart gebliebenen M~nnchens. Variation 4, die Doppelstrophe, wechselt im Prinzip ebenso, wie in Fig. 3. Aul]erdem zeigt sieh, dab Variation 4 gegeniiber 1 und 2 und 3, den normal langen GesEngen, negativ korreliert.

Heft 3] 1967 ] Gesa~ng des Fitis 277
Die Variationen 1--3 sind in den Morgenstunden am st~irksten vertreten (bei grSflter Gesangs~tivit£t), fallen dann zu einem Tiefpunkt in der 11. Stunde ab (niedrigste Aktivit~it des Individuums) und nehmen sehliel31ich nicht in dem MaBe zu, wie es nach der in Fig. 3 dargestellten Aktivit£tskurve zu erwarten w£re. Na~ dem Minimum zwischen 16 und 17 Uhr steigen sie dann bis zum Aktivit£tsende an, erreiehen abet nicht die Werte des Vormittags. Variation 2 zeigt einen £hnlichen Verlauf wie die Kurve der Gesangs- aktivitiit (vgl. Fig. 5).
Fig. 6 stellt die Wi~derholungsgr5i3e im Tagesgang dar. Sie liegt im allgemeinen zwischen 7,6 und 22,6 °/0. Bis znr 8. Stunde bleibt sie nahezu gleida und verh~lt sich mit kleinen Schwankungen ebenso wie die Gesangsaktivit~it (s. Fig. 1), W~ihrend je~e bis zum friihen Nachmitag wetiterhin abnimmt, singen die Fitisse nun wieder zu- nehmend einfSrmig bis zur 16. oder 17. Stunde. Hier tritt ein deutlieher Sprung ein: Der Gesang wird wieder variabler, um zum Abend wiederum einfSrmiger zu werden. Ab 10 bis 11 Uhr liegt also ein g~inzlida andersartiger Verlauf als bei der Gesangsaktivitgt vor, der einer n e g a t i v e n Korrelation entspreehen wfirde. M~nnchen 5 sang dagegen nachmittags und abends auffallend stereotyp (gestrichelte Linie) und somit im entspre- chenden Verh£1tnis zum Aktivit~tsniveau. Dieses Verhalten hebt sich auffaUend yon allen anderen Individuen ab und deutet die groBe individuelle Variationsbreite in der Bezie- hung zwischen Aktivit~t und Wiederholungsh£ufigkeit an.
4. Variationen in der Strophenliinge
Auch die Gesangsl£nge ist tageszeitlichen Schwankungen unterworfen. So wur- den 171 am 2. Mai yon drei M~innchen (Schwarz: 39, ~ 5: 63, WeiBgelb: 69) zu verschiedenen Tageszeiten aufgenommene Ges~inge auf ihre L~nge gepriift.
Es zeigte sich, da~ alle drei im Laufe des Tages zunehmend l£nger singen. Das Minimum liegt jeweils in den friihen Morgenstunden (2,66--2,97 see.). Darauf folgt ein stetiger Anstieg, der zu Beginn der zweiten Hauptgesangsphase gegen 17.00 gipfelt. (3,17--4,46 sec.) Bis zur 19. Stunde werden die Ges~inge wiederum verkiirzt his auf Mittelwerte yon 2,70--3,74 see.
]~. Modifikation w~ihrend des: Revierkampfes
Dringt ein Rivale in das Revier eines M~nnchens ein, wird der Gesang, sobald der Eindringling als solchcr erkannt wurde, fiir eine gewisse Zeit g£nzlich unter- brochen. Der Revierinhaber lauscht w£hrenddessen dem Gesang des Eindringlings und verfolgt dessen Bewegungen. Mehrere Strophen des fremden Vogels sind not- wendig, um das Angriffsverhalten auszulSsen. Dann folgen Suchflfige, begleitet yon sirrenden Lauten (Abb. 4 F) und einem dcm Erregungsruf £hnlichen ,,hi~d" (Abb. 4 I), kreischende Laute (Abb. 4 H) und Rufe wie ,,hid-jek-}ek" (Abb. 4 D--E), ,,wing flapping" (MAY, 1949) und gelegentlich Imponierfliige. Dcr Gesang ist vor- erst vollstiindig gehemmt. Allmiihlich und meist crst, nachdem der Rivale in Rich- tung Reviergrenze oder dariiber hinaus getrieben wurde, 15st sich die Hemmung. Nun werden Gesangsbruchstiicke unmittelbar zwischen den verschiedenen Rufen oft sehr laut und mit deutlicher Betonung dcr einzelnen Elcmente herausgeschleu- dert. Diese Hemmungsphase ist individuell und situationsgem~il3 yon unterschied-
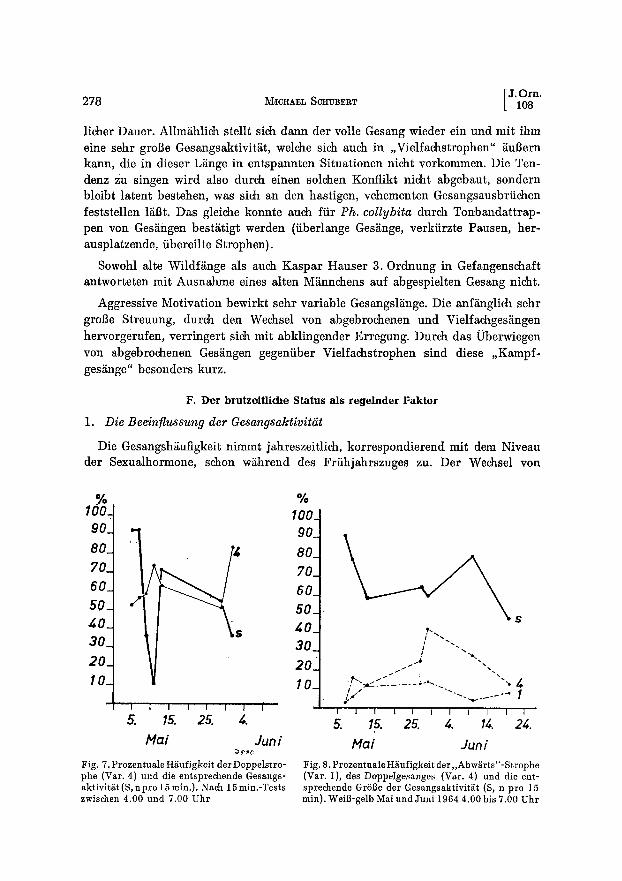
rJ.o . 278 M IcaA~,. Scn~Bv, aT [ 108
licher Dauer. Allm~ihlich stellt sich dann der volle Gesang wieder ein und mit ihm eine sehr groBe Gesangsaktivit~it, welche sich auch in ,,Vielfachstrophen" ~uBern kann, die in dieser L~nge in entspannten Situationen nicht vorkommen. Die Ten- denz Zu singen wird also durch einen solchen Konflikt nicht abgebaut, sondern bleibt latent bestehen, was sich an den hastigen, vehementen Gesangsausbriichen feststellen l~Bt. Das gleiche konnte auch ffir Ph. collybita durch Tonbandattrap- pen von Ges~ingen best~tigt werden (iiberlange Ges~nge, verkfirzte Pausen, her- ausplatzende, fibereilte Strophen).
Sowohl alte Wildf~nge als auch Kaspar Hauser 3. Ordnung in Gefangenschaft antworteten mit Ausnahme eines alten M~nnchens auf abgespielten Gesang nicht.
Aggressive Motivation bewirkt sehr variable Gesangsl~inge. Die anf~inglich sehr groBe Streuung, durch den Wechsel yon abgebrochenen und Vielfachges~ngen hervorgerufen, verringert sich mit abklingender Erregung. Dutch das fJberwiegen von abgebrochenen Ges~ingen gegeniiber Vielfachstrophen sind diese ,,Kampf- gesiinge" besonders kurz.
F. Der brutzeitliche Status als regelnder Faktor
1. Die Beeinflussung der Gesangsaktivitiit
Die Gesangsh~ufigkeit nimmt jahreszeitlich, korresportdierend mit dem Niveau der Sexualhormone, schon w~hrend des Frfihjahrszuges zu. Der Wechsel yon
,# 90_
86
76
66
50_ 40_ 30_
20_
10_
%
100_ " 90
'~ 80 70_ 6O 502 ,~0
s 30 I
20 J 10_
I t ~ l a J I i
5. 1£ 25. ~.
Mai Jun i
Fig. 7. Prozentuale Hi~ufigkeit derDoppelstro- phe (Var. 4) und die entsprechende Gesangs- aktivit~t (S, n pro 15 rain.). Nach 15 min.-Tests zwischen 4.00 und 7.00 Uhr
s
!
/ t ~ ~x
I I I ! I I I I I I I
5. 15. 25. & l& 24.
Mu i Jun i Fig. 8. Prozentuale H/iufigkeit der,,Abw~irts"-Strophe (Vat. I), des Doppelgesanges (Vat. 4) und die ent ~ sprechende GrSBe der Gesangsaktivit~t (S, n pro 15 rain). WeiB-gelb Mai und Juni 1964 4.00 bis 7.00 Uhr

Hle.ft73 ] Gesang des Fitis 279
8ubsong zum Reviergesang hat sich, wenn die Fitisse bei uns eintreffen, bereits vollzogen. Aueh die nordisehen Durehzfigler singen, wenn aueh unregelm~iBig und meist nur am friihen Morgen, vollen Motivgesang. Die Gesangsrate steigt mit der Griindung des Territoriums sprunghaft an. Aueh MAY (1949) fand, dab mit der Benutzung der Naehbargebiete die Gesangshgufigkeit zunimmt. Das Ansingen stei- gert und synehronisiert die Gesangsrate, es ist im April am h~ufigsten. Mit dem Eintreffen der Weibehen in den letzten April-Tagen erreieht die Gesangsinten- sitiit ihren HShepunkt, doch in den Tagen der Balz tritt ein kurzfristiger Riick- gang ein (Fig. 7), w~ihrend die unverpaart gebliebenen M~nnchen bis in den Juli hinein noch regelm~i~ig singen (Fig. 8) (s. auch DAws, 1958, CURm, 1959, v. HAARrMAN, 1956). So sang das seit dem 28'. oder 29. April verpaarte ~ 5 be- reits am 29.4. auffallend weniger als vorher. Der stufenweise erfolgende Riick- gang den das Diagramm von Cox (1944) nahelegt, diirfte vor allem unverpaarte Stficke betreffen, da die verpaarten nach eigenen Feststellungen ihre Gesangsrate im Mai wieder etwas steigern, dabei jedoch nicht die Ausdauer der ledigen M~inn- chen erreichen.
Mit dem Einsetzen der Vollmauser Ende Juni bis Mitte Juli tritt Ruhe ein. Nur selten hSrt man noeh hier und da einzelne Strophen. Naeh der Mauser (Anfang August) kann die Aktivit~t bei vielen H~hnen fiir 2--3 Wochen noch einmal auf- leben. Sie singen sich dabei oftmals regelrecht an, imponieren, attackieren und zeigen iiberhaupt ein ausgepr~gtes Revierverhalten. Mehr als 90 Strophen je 15 min. sind keine Seltenheit. In manchen Gegenden hSrt man allerdings zu dieser Zeit kaum noch einen Vogel singen, obwohl anderorts (an bestimmten Tagen) starke Ansammlungen zu verzeiehnen sind. M5glieherweise liegt die grSl~ere terri- toriale Aktivit~it bei den Durchziiglern. PALMGREN (1934) nennt Ph. trochilus einen der fleiBigsten Herbstsiinger in Finnland, PALM (1951) nimmt an, dab bei Ph. col- lybita die JungvSgel, die friiher als die Alten ziehen, aktiver singen.
2. Unterschiede in der Hiiufigkeit von Variationen
Verpaarte M~nnchen singen im allgemeinen variabler als ledige. So wurden bei Blau und ~ 5 (beide verpaart) 37 resp. 41 unterschiedliche Gesangstypen fest- gestellt, bei WeiB-gelb (Sp~tbriiter) und Schwarz (unverpaart) nut 20 resp. 24. Somit entfallen auf einen ,,neuen" Gesangstyp x Wiederholungen: Blau: 4,49 (n -- 166); ~ 5 :4 ,58 (n -- 197); WeiB-gelb: 9,80 (n = 196); Schwarz: 12,25 (n = 294).
Die Junggesellen trugen einige Ges~inge sehr dauerhaft vor, z. B. Schwarz den Typ 2/4 (Abb. 3 A--C), WeiB-rot u. a. 10/39 (Abb. 2 D) und WeiB-griin den Typ 7/9 (Abb. 2 C), wohingegen die verpaarten Miinnchen schneller die Motive wech- seln. So wurden nur 7 der 41 Typen yon ~ 5 an mehr als einem Tage festge- stellt. Motive aus der Zeit der Revierbesetzung halten sich jedoch bei beiden am l~ngsten gegenfiber den Mai- und Juni-Ges~ngen (Fig. 9).

280 ~ICHAEL SCHUBERT J.Orn.
108
6 5 . blau
J u n i
• 10.
2 6 .
Mai
13. 9.
: 5. • 2 , I , , " ~ -
25.
- - - 20.
s c h w a r z ,w. - g r ~ n, w. - r o t, w. : ge lb
i
I
m
. . . . . . . . . . . =Ap r . . . . . . . . . . . . .
50 ~0 30 20 10 10 20 30 ,~0 50 Fig. 9. Erstmaliges Erscheinen und Dauer yon Gesangstypen. Links verpaarte, rechts unverpaarte Die L~nge der Siiulen gibt an, wie lange die an den betreffenden Tagen erstmals aufgenommenen
Gesangstypen weiterhin gesungen wurden
Typisch ffir die Zeit der Revierbesetzung und -verteidigung ist eine hohe Ge- sangsrate bei relativ kurzem und lautem Gesang, wi~hrend schon vom 5. bis 15. Mai an Tendenzen zum Ruhegesang (Subsong) feststellbar sind: kurzfristige Motive, sporadischer, leiser und wenig priignanter Vorgesang, Doppelstrophen, Einleitungslaute, niedrige Gesangsaktivit~t*).
In Fig. 5, 7 und 8 wurden die gemittelte Gesangsaktivit~t und verschiedene Variationstypen nach 15 min.-Tests ausgewertet. Es wurden Z~hlungen eines ver- paarten (Blau) und zweier lediger (WeiB-gelb und Schwarz), die yon 4 bis 7 Uhr gemacht wurden, yon Anfang Mai bis Ende Juni verglichen. Bei allen drei H~hnen ist zu bemerken, da~ Anfang Mai die Gesangsaktivitiit deutlich abnimmt, am deut- lichsten bei Blau, der am 10.5. besonders intensiv balzte. Schon am 13.5. war er wieder h~ufiger zu hSren, darauf f~llt die Kurve wiederum ab. Auch .die iibrigen zwei sangen um den 10. Mai nicht sehr intensiv, s t e i g e r t e n sich aber im weiteren Verlauf des Mai. Doppelges~inge traten bei WeiB-gelb, Blau und ~ 5 erstmals am 2.5. auf, bei Schwarz am 7.5. und bei Rot am 9.5. Doppelstrophen sind typiseher Ausdruck der epigamen Komponente im Gesang des Fitis. Bei Blau erseheinen sie in der Balzzeit (10.5.) am h~ufigsten, bei Weil3-gelb Ende Mai, bei ledigen Miinnchen (Schwarz, WeiB-griin) dagegen weitaus seltener. Inwiefern die Gesangsaktivit~t im Verlauf der Brutzeit die H~ufigkeit der Doppelstrophen be- riihrt, ist nicht mit Sicherheit festzustellen.
Doppelstrophen kSnnen, wie bereits festgestellt wurde, aueh im Revierkampf auftreten. Hier scheinen zwei Mofivationen wirksam zu sein: die Tendenz, den
*) A~MSVRO~G (1963) erw~gt, ob es eine positive Beziehung zwischen der Anzahl artgleicher Reviernachbarn und dem Umfang der Variabilit~t gibt. Zur ErSrterung dieser Frage reicht das vorliegende Material nicht aus.

Heft 3] 1967 J Gesang des Fitis 281
singenden Rivalen nicht ,,zu Wort kommen zu lassen" (auch bei Ph. collybita beobachtet) und die enthemmte, angestaute Sing-Reaktion. Ganz anders als diese klingen die Doppelgestinge in der Balzzeit und gerade vor der Kopula: leiser, weicher, schwatzhafter Vortrag, hiiufiges Sirren und keine aggressiven Laute.
Auf das ,,Leiern" einiger M~innchen wurde bereits oben eingegangen. Diese besonders ffir Blau und (~ 5 bezeichnende Variante stellte sich bei beiden zuneh- mend h~ufiger in und nach der Balzphase ein, so dab die meisten Gesiinge bereits Ende Mai davon ergriffen waren.
Auch die Tendenz zur Verl~ngerung der Phrasen wird mit fortsehreitender Brutzeit deutlich. Ein Motiv yon ~ 5 endete am 20.4. beispielsweise mit einem V-fSrmigen Schluglaut, am 29.4. dagegen mit V V. Eine Endphase yon Sehwarz bestand seit dem I 1.5. vorwiegend aus 4--5 start 1--3 Elementen (Abb. 3 A u. C). Seit dem 27.5. sang Weil3-gelb die harten Sehlgge einer Variation (Var. 2) doppelt.
Nicht selten werden sp~ter neuartige Elemente in die hestehenden Phasen eingegliedert, wodureh die Strophe, falls nieht andere Elemente daffir verschwin- den, liinger wird. Seit dem 26.5. sang Sehwarz viele Gesgnge in einem neuartigen Crescendo.
Wenn die Zeit der heftigsten Revierk/impfe vorfiber ist (Mitte Mai) wird der Vortrag bei vielen d leiser, weieher und lockerer (vgl. MAY [1949], der feststellt, der Gesang sei ab 10. 5. ,,more softly"). Das Leiern von Blau verflachte sich Mitte Mai so, dab es die Intervalle verlor und in ein einfaches ,,Dudeln" fiberging.
Seit dem 10.6. verlor sich die Strophigkeit der Gesiinge von Schwarz mehr und mehr, .die Pausen zwischen den Gesgngen wurden sehr kurz, daffir wurden hgufiger lange Generalpausen eingelegt (charakteristische Merkmale des Subsongs). WeiB-rot sang am 16.7. sehr variabel und manche Partien klangen neuartig, obwohl er grSBtenteils alte Grundmotive sang (6 Ges. in 3 neuen, 18. Ges. in 3 alten Typen).
3. Die Hiiufigkeit von ,,Sirren" und ,,sit"-Lauten
Mitte Mai bis Ende Juni hSrt man von Fitissen besonders hi/ufig einen sirren- den Laut, weleher meistens ,/on (~ ge~ugert wird, die im Leerlauf imponieren (Abb. 4 C). Er wurde zuletzt am 27.6. gehSrt, danach erst wieder naeh der Som- mermauser ab Anfang August. Auch die JungvSgel ,,seheinbalzen" bereits zu dieser Zeit. MAY (1949) stellte Sirren in der spgteren Fortpflanzungsphase am hgufigsten lest (ab Ende Mai zunehmend hgufig bis zur 2. Woche im Juni) u~.d erw~igt, ob es einen Ersatz ffir den Gesang darstellt.
Ein anderer Ruf ist im Gegensatz zum Sirren obligatorisch mit Gesang gekop- pelt, das ,,sit", ein sehr leiser hoher Laut, der 1--4-, meist aber 2--3mal in Abstiinden von ca. 1 see. vor einer Strophe ge~iuBert wird. Er tritt individuell verschieden Miufig auf und ist nur von Ende Mai bis Anfang Juli zu hSren. Er ist charakteristiseh fiir den spiiten Motivgesang und ist wahrseheinlieh den ,,trett"-Rufen yon Ph. collybita homolog.
[J.Om. 282 MICHAEL SCHCBERV [ 108
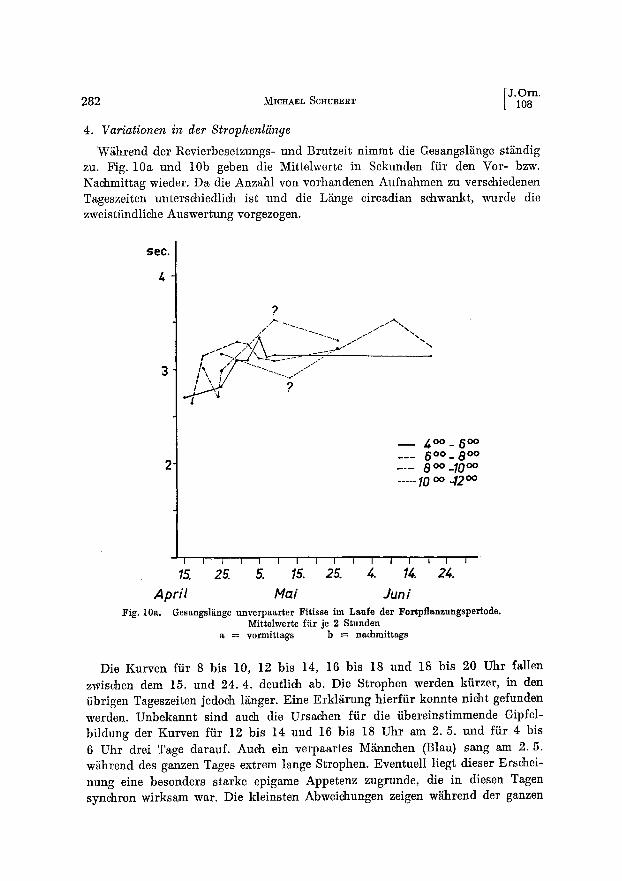
4. Variationen in der Strophenliinge
W/ihrend tier Revierbesetzungs- und Brutzeit nimmt die Gesangsl~nge st~ndig zu. Fig. 10a und 10b geben die Mittelwerte in Sekunden ffir den Vor- bzw. Nachmittag wieder. Da die Anzahl yon vorhandenen Aufnahmen zu verschiedenen Tageszeiten unterschiedlich ist und die L~nge circadian schwankt, wurde die zweistiindliche Auswertung vorgezogen.
s e c .
3
A p r i l Fig. lOa.
?
/'.,. f / .............. ....- I
4 0 0 _ 6 ° o ___ 6 o o _ 8 0 0 - . - 8oo_10oo ..... 10oo 42 °o
i l J I I t t I t t i i I t J !
15. 25. 5. 15. 25. ~. 1~. 2~.
I ~a i Jun i G e s a n g s l E n g e u n v e r p a a r t e r F i t i s s e i m L a u f e d e r F o r t p f l a n z u n g s p e r i o d e .
Mittelwertc ffir je 2 Stunden a -- vormi t tags b ~ nachmit tags
Die Kurven fiir 8 bis 10, 12 bis 14, 16 bis 18 und 18 bis 20 Uhr fallen zwischen dem 15. und 24.4. deutlich ab. Die Strophen werden kfirzer, in den iibrigen Tageszeiten jedoch l~nger. Eine Erkliirung hierffir konnte nicht gefunden werden. Unbekannt sind auch die Ursachen fiir die iibereinstimmende Gipfel- bildung der Kurven fiir 12 bis 14 und 16 bis 18 Uhr am 2.5. und fiir 4 bis 6 Uhr drei Tage darauf. Auch ein verpaartes M~innchen (Blau) sang am 2.5. w~hrend des ganzen Tages extrem lange Strophen. Eventuell liegt dieser Erschei- nung eine besonders starke epigame Appetenz zugrunde, die in diesen Tagen synchron wirksam war. Die kleinsten Abweichungen zeigen w~hrend der ganzen
Heft 3] 1967 J Gesang des Fitis 283
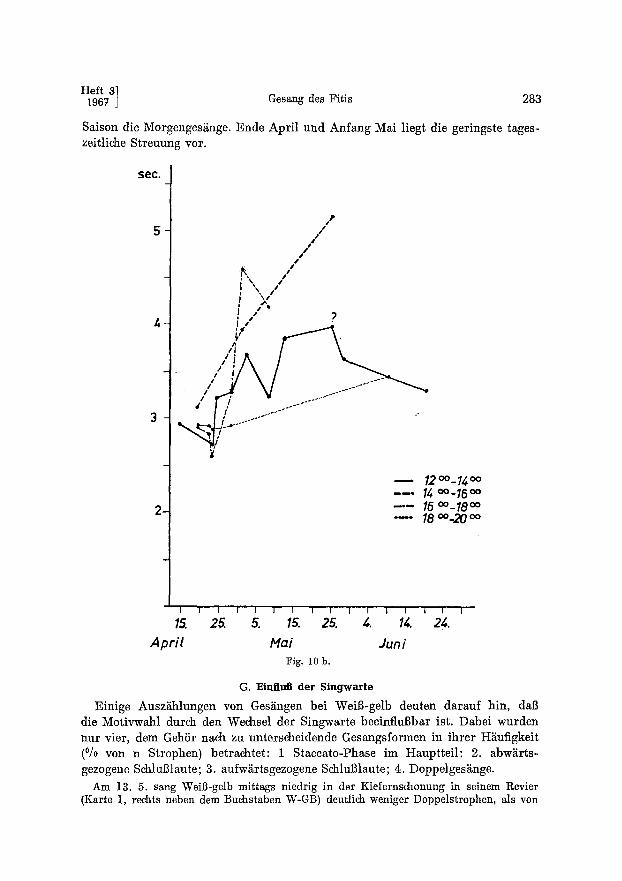
Saison die Morgenges~nge. Ende April und Anfang Mai liegt die geringste tages- zeitliche Streuung vor.
s e c .
5
.
/ #g g
SS
,,4" . / ] A /
12 oo_ I~ OO - - . 1 4 o o - 1 6 o o
- ' - 15 ° ° - 1 8 ° ° . - - . 18oo_20oo
I I I I I I I I | I I I I | t
15 25. 5. 15. 25. & l& 2.4.
Apr i l IVlai Jun i Fig. 10 b.
G. Einflu~ der Singwarte
Einige Ausz~hlungen yon Ges~ngen bei Weil3-gelb deuten darauf hin, dal3 die Motivwahl durch den Wechsel der Singwarte beeinfluBbar ist. Dabei wurden nur vier, dem GehSr nach zu unterscheidende Gesangsformen in ihrer H~iufigkeit (°/o yon n Strophen) betrachtet: 1 Staccato-Phase im Hauptteil; 2. abw~rts- gezogene Schlul31aute; 3. aufw~rtsgezogene SchluBlaute; 4. Doppelges~nge.
Am 13. 5. sang Weil~-gelb mitt~gs niedrig in der Kiefernschonung in seinem Revier (Karte l, rechts neben dem Buchstaben W-GB) deutlich weniger Doppelstrophen, als yon

284 ~/[ICHAEL SCHUBERT [J.O~n.
oiner froistehenden jungen Birke aus ca. 5 m H5he in dem gleichen Gebiet. Am 27. 5. ab 5.21 Uhr brachte er in den hohen Erlen (NE yon den Buchstaben W-GB) fast ausschlieB- lich Doppelgesang, unmittelbar vorher von einer anderen Singwart8 jedoch innerhalb yon 14 min nur 3 real.
Am 10.6. wurde derselbe Vogel yon 5.45 bis 6.00 Uhr verhSrt. In den hohen Erlen (s. o.) sang er 24 Strophen, davon 0 °/o in Var. 1; 75,86 °/o in Var. 2; 10,35 °/0 in Var. 3 und 13,79 °/o in Var. 4. Unmittelbar vorher sang er in der Kiefernschonung 41 Strophen: 2,44 °/o in Var. 1; 53,66 °/o in Vat. 2; 43,90O/o in Var. 4.
Als die Jungen von ,,WeiB-gelb" am 20.6. zwei Tage alt waren, sang er sehr h~iufig im siidliehen Tell des Reviers in hohen Birken am Rando des Hoehwaldes unmittelbar in Nestn~ihe aus 12--15 ra Hiihe und aus 3--5 m HShe in der Kiefernsehonung. 16.15 bis 16.19 Uhr und 19.37 bis 19.41 Uhr brachte er in letzteremGebiet 14 ~-21 Strophen, davon 14,2 °/o in Var. 1; 48,57 °/o in Var. 2; 2,86 °/o in Vat. 3 und 34,280/o in Var. 4. 16.19 bis 16.30 Uhr, 19.30 b~s 19.37 Uhr und 19.41 bis 19.44 Uhr wurdc er hoch fiber dem Nest verhSrt: 40 ~- 14 ~ 13 Strophen, davon 10,45°/o in Vat. 1; 67,16 °/o in Var. 2; 16,42 °/o in Var. 3 und 8,98 °/o in Var. 4.
Somit sang dieser Hahn Variation 2 unabhiingig yon der Lage der Singwarte im Revier in hohen Biiumen h~iufiger als in den niedrigen Kiefern und Birken des Gebietes, seltener dagegen die Motive 1 und 3. Nach ARMSTRONG (1963) singt ein Vogel, dessen Reviereifer nachl~£Bt, yon einer niedrigeren Singwarte. Auch bei Ph. trochilus halten sich singende Exemplare vorwiegend in den Baum- kronen auf, w~ihrend sieh Balz, Begattung und Leerlauf-Imponieren ve t allem in niedrigen B~iumen und Biischen abslaielen. In Ubereinstimmung mit MAY (1949) wurde beobachtet, dab die Fitisse am h~iufigsten hohe Singwarten benutzen, die mehrere Minuten durchgehend singen und dann etwa die gleiche Zeitdauer zum Fressen ben6tigen gegeniiber anderen, die nur einen Moment singen und dann wieder auf Nahrungssuche gehen. DAws (1958) beobachtete, dab Pipilo erythrophthalmus mit fortschreitender Brutzeit in stetig abnchmender H6he singt (so auch ein unverpaartes c~). Monticola saxatilis hat bis zu 2 km entfernt liegende gesonderte Singpliitze fiir Revier- und entspannten Gesang (FARKAS 1955)*). Nach KOFFA~ (zit. nach ARMSTRONG 1963) variiert der Gesang der Heidelerche , , . . . according to circumstances and the mode of utterance. The nearer the nest and the female, the softer the song."
H. D i s k u s s i o n
Der Fitislaubsi£nger ziihlt neben den asiatisehen Arten Ph. proregulus (besonders die Ssp. chloronotus), subviridis, nitidus und magnirostris zu den am abweehs- lungsreichsten singenden Laubsiingern. MARLV.R & ISAAC (1960) fanden bei 47 um Cambridge aufgenommenen Strophen mindestens 12 verschiedene Themen. Vier willkiirlich gew~hlte Ges~inge yon 4 ~ c~ unseres Gebietes batten beispiels- weise 25 versehiedene Elemente. Wenn HARTSnORNE (1956) angibt, Ph. trochilus w~re ein ,,extremely non-versatile singer", so liegt hier sicher eine Verwechslung mit Ph. collybita vor.
*) Grallina cyanoleuca singt vom Nest aus ein Gesangselement mit besonderem Vibrato (RosiNSO~ 1949).

t-teft 3] 1967 I Gesang des Fit is 285
Die hier vorge.nommene Differenzierung des Repertoires in Grundtypen crwies sich als Notwendigke.it, um die innerhalb einer Brutsaison anfallenden Variationen ann/ihernd fiberschauen zu kSnnen. Innerhalb jeden Typs liegen wiederum Varia- tionen in der Anzahl, L~nge und Feingestaltung der Elemente, der Lautst~irke und Betonung vor, welche allerdings in diesem Rahme.n kaum be.riicksichtigt wer- den konnten.
In einem entspannten, stSrungsarmen Umfeld ist das Spektrum von Variations- mSglichkeiten besonders grol], das Verhalten ersche.int, zumal in bestimmten juvenilen Stadien, abwechslungsre.ich, locker, oft spielerisch und ,,funktionslos". Der Gesang des Fitislaubs~ngers (wie auch mancher anderer Passeres) zeigt diese Tendenz nicht nur bei JungvSge.ln im Herbst und Winter und in der allj~ihrliehen Reifung des Reviergesanges der Altv5gel im Friihjahr (Sekund~rgesang, Subsong), sondern auch in der Fortpflanzungsphase.
Laubs~nge.r sind als visue.ll unauff~llige Baum- und BusehvSgel insbesondere auf akustische. Kommunikation angewiesen. Aul3erhalb der Brutzeit ist beim Fitis (mit Ausnahme. der Sp~tsommerbalz) diese Notwendigkeit nieht mehr gege.- ben, da e.r wie die meisten Sylviinae ein ausgesprochene.r Einzelg~nger ist. Er verffigt weder fiber Stimmffihlungs- noch Lockrufe (das ,,hiii", Abb. 4 F und G, ist le.diglich e.in Erre.gungsruf). Mit Ende August klang bei den adulten Wild- fiingen die Singbereitsehaft ab und trat erst wieder im Februar und M~rz, naeh beendeter Vollmauser, verst~irkt ein. WildvSgel sind im Herbst offenbar stimm- freudiger; HOESCH und NXE~A~IER (1940) beobachteten in Siidwestafrika vie.lfach singende Exemplare im Oktober. Die e.rste.n Frfihjahrsges~nge unserer K/ifigvSgel trugen zu dieser Zeit den Charakter yon Sekundiirges~ngen (Subsong): leise, sehr abwechslungsreich, kurze Zwisehenpausen, lange Generalpausen, erhShte, sehr variable Gesangsl~nge, Einleitungslaute meistens in Form yon Erregungsrufen und Imponierlauten (,,Sirren"). Dieser sich allj~ihrlich +- ausffihrlich wieder- holende Reifungsprozel~ der informationssendenden Signalstrukturen findet seinen (vorl~ufigen) AbschluB mit de.m Entstehen der ffir den Empf~nger optimalen, pr~gnanten artspe.zifisehen Signalstruktur, des Motivgesanges (Prim~ir- oder Re- vierge.sang). In zunehmendem MaBe wird ein Gestaltungsdruck wirksam, durch den das Repertoire insgesamt gerafft und konzentriert wird (stetiger Vortrag, bis zu 105 Strophen je 15 rain., grSBere Amplitude, zune.hmende Phasierung, Einengung des Variationsspe.ktrums, kurze akzentuierte Ges~inge unter Fortfall yon Einleitungsrufen und einer zweiten Strophe). Diese Kriterien folgern ins- besondere aus den Beobachtungen und Analyse.n der (vornehmlich territorial motivierten) ,,April-Ges~inge". Die Gesangsthemen sind relativ dauerhaft und zu allen Tageszeiten zu hSren. Sie bleiben fiber mehrere Tage oder Woehen kaum modifizie.rt erhalten, am l~ngsten bei unverpaarten ~. Doppe.lges~nge sind vorerst selten. Die. Strophen klingen pr/ignant und besitzen keine verflachten Phrasen (ira Vergleich zu sp/iter aufkomme.nde.n Themen).
Blau (verpaart) sang nach der Balz nur ein einziges yon 15 Themen weiter, Schwarz ffihrte indessen 5 yon 17 und WeiB-grfin {heide unverpaart) 3 yon 11 ,,April-Motiven"

I J" Orn. 286 M~eHAEL SCHUBERT [ 108
im Mai und Juni fort. Es konnten in keinem Falle fiir aggressiven oder fiir Balzgesang spezifische Themen gefunden werden.
HOMANN (1960) und HO~A~N & GWINNER (1963) unterscheiden bei Ph. collybita Revier-, Rivalen-, Erregungs-, Balz- und Werbegesang. Nach eigenen Beobachtungen sind sie alle durch Oberg~nge verbunden und ihre Schwellen liegen individuell sehr unterschiedlich. Wiihrend der Balzgesang fehlt, sollen im Herbst Erregungs-, Rivalen- und gelegentlich Werbegesang vorgetragen werden.
~nderung der Motivation zeigt sich nur in der Art des Vortrages: Balzgesang ist oftmals yon Imponierlauten begleitet, und die einzelnen Ges~nge sind durch lange Pausen getrennt; bei verminderter Gesangsaktivit~it sind die Ges~nge laug, es gibt viele Doppelges~inge, die Lautst~rke ist geringer als beim Reviergesang (aber grS~er als beim Subsong), und es treten zahlreiche neue, kurzfristige Motive auf. Wir mSchten diese Form des Werbegesanges als Ausdruck des verminderten Gestal~druckes betrachten. Er ist in der Zeit der Revierbesetzung und -Verteidi- gung am st~rksten wirksam, w~hrend der Gesang ffir das ~ nach der Verpaarung praktisch bedeutungslos wird. Dem ~ kommen bei der Aufzucht keine Aufgaben zu, alleinstehende Weibchen kSnnen mit dem gleichen Erfolg ihre Jungen auf- ziehen wie solche, die bei der Ffitterung vom ~ unterstfitzt werden. Auch die Revierverteidigung verliert mit fortschreitender Brutzeit mehr und mehr an Be- deutung, da die Reviergrenzen fixiert sind, der Durchzug beendet ist und unver- paarte M~innchen in der Regel ebenfalls reviertreu sind. (Oftmals reagieren Brut- vSgel Ende Mai und im Juni auf abgespielten Gesang nicht mehr aggressiv, son- dern schenken der Attrappe keine Beachtung.) Das wichtigste Kommunikations- mittel der Brutzeit ffir die Partner untereinander wird der Erregungsruf ,,hi~i", der nunmehr als Warnruf ffir StSrungen in Nestn~ihe fungiert.
In der Literatur findet man fiber die Einwirkung des Fortpflanzungsstadiums auf die Gesangsqualit~t nur wenige, teils widerspriichliche Hinweise. Nach S c~z (1950) haben Grfine Lanbsiinger in der sp~teren Brutzeit (selten) zwischen den Strophen ein ,,leises wohlklingendes Schw~tzeln" und einen hohen trillernden Ruff Nach Verlust der Brut Mitte Juli sang ein c~ dichtend und mit halblauter Stimme eine Gesangsform, die noch Elemente des Motivgesanges enthielt und deren Er- scheinen auf die abnehmende Gesangsaktivit~t hindeutet.
DE~Ius (1963) beobachtete, da~ die Ges~nge von Alauda arvensis w~ihrend der Paarungszeit extrem lang und w~hrend der ersten Brut am kiirzesten sind. Die H~iufigkeit der Singflfige nimmt w~hrend der Paarungszeit ab, steigt dann bis ca. 8. Mai an, sinkt wiederum bis zum 22.5. (Brutphase), nimmt wieder bis ca. 4.6. zu und f~llt schlieBlich sehr stark ab.
Nach SAUER (1955 a) ist der Balzgesang der MSnchsgrasmiid~e (Sylvia atrica- piUa) reich an Imitationen und verhiiltnism~Big arm an lauten Elementen und kann bei Erscheinen eines c~ im Revier in vollen lauten Motivgesang umschlagen. Dorngrasmficken (Sylvia communis) singen nach der Revierbesetzung sehr va- riationsreich, solange weder Revierk~impfe noch Balz stattfinden. Sie verkiirzen und vereinheitlichen die Motive w~hrend der Balzzeit etwas, doch werden sie als-

Hle9I•73] Gesang des Fitis 287
bald wieder umfangreicher. Atternierend singende Naehbarn haben sehr formali- sierten, kiirzeren Gesang (so auch atricapilla). Dorngrasmiicken haben den grSB- ten Melodienreichtum im Herbst, Winter und Friihjahr. Sie singen in der Paa- rungszeit ,,verbalzt" auf und ab pendelnd. Der Vortrag ~ndert sich jahres- und tageszeitlich bei allen einheimischen Grasmficken (SAVER 1955 b). Troglodytes aedon singt, sobald ein Weibchen eingetroffen ist, energischer und wechselt nach der Paarbildung zu einem weichen, tieferen, leisen Nistgesang (ARMSTRON~ 1963). Nach GusE (1964) (1964, unverSffentlichte Staatsexamensarbeit an der math. nat. Fakult~it der Humboldt-Universitgt Berlin) ist die Anzahl unterscheid- barer Laute von Amsel (Turdus merula) und Singdrossel (Turdus philomelos) im Frfihjahr grSBer als spgter im Jahr. Bei Pipilo erythrophthalmus dagegen werden das Repertoire und das stimmliche Volumen mit ausklingender Brutsaison kleiner (DAvis 1958). Gartenrotschw~nze (Phoenicurus phoenieurus) sollen w~hrend des Nestbaues besonders zu Imitationen neigen (BuxTON 1950). Trauerschngpper (Ficedula hypoleuca) iindern mit vorrfickender Brutzeit die Motive, die Pausen zwisehen den Ges~ngen werden l~nger, die Strophen k ii r z e r. Ein drei- oder viersilbiges Motiv tritt bei flauer Gesangsstimmung mittags und nach der Paa- rungszeit auf (CuRIO 1959). NeuntSter (Lanius collurio) lassen mit dem Gesang bald nach dem Erscheinen eines Weibchens naeh und reduzieren die H~ufigkeit ihrer harten Rufe. Die ,,ty-Strophe" der Goldammer (Emberiza citrinella) ist im Juli und insbesondere im Herbst h~iufiger. KAISER (1965) wertet sie als eine ,,Er- scheinung des Abflauens" der Gesangsaktivitiit. Die Anwesenheit des ~ wirkt bei Buehfinken nicht nur auf die Gesangsaktivit~t hemmend, sondern es stellen sich w~ihrend der Balz auch kurze gepreBte Variationen mit oft wiederholten End- phrasen ein (PouLSEN 1958). DAVIS (1958) beobachtete Gesangshemmung nach dem Schliipfen der Jungen und fiihrt diese auf die Ausschiittung des Hormons Prolaetin zurfiek, welches das Aufzucht-Verhalten aktiviert. Nach MESSMER'S (1956) ist der Balzgesang der Amsel dem Subsong i~hnlich. ARMSTRONG (1963) stellt fest, dab w~hrend der Reviergriindung Zunahme der Lautst~irke, der Per- sistenz und der V a r i a t i o n s b r e i t e stattfindet, w~ihrend ,,courtship and nuptial songs" zu einer w e i e h e r e n und w e n i g e r v a r i a b l e n Vor- tragsweise neigen. Diese Modifikationen diirften im wesentlichen, korrespondierend mit der Regelung der Gesangsaktivit~it im Jahresverlauf, durch die Ausschiittung von Gonaden- und/oder gonadotropen Hormonen gesteuert werden.
Eigentiimlicherweise trifft man bei der Untersuchung der Qualitgt yon Vogel- gesgngen im Tagesverlauf auf Tendenzen, die sich z.T. mit den jahreszeitlichen und brutzeitlichen Modifikationen vergleichen lassen.
Schon CRAm (1943) marhte die erstaunliche Feststellung, dab Myiarchus virens (Tyran- nidae) fiber einen besonderen Tagesanbruch- oder Zwielicht-Gesang verffigt. Er beginnt ca. 1 Stunde vor Sonnenaufgang und dauert etwa 30 rain. BOR~OR (1961) bemerkt dazu, dal3 diese Gesangsformen ,,are undoubtedly influenced by light intensity, but their significance to the birds is not clearly understood". Er stellt ferner fest, dab die meisten VSgel am friihen Morgen zusammenh~ingender als in der Tagesmitte singen. Auch Musci- vora tyrannus (Tyrannidae) solt nach N~cE (1931) einen speziellen Morgengesang vor-

[J.Orn. 288 ~iCHAI~L SCHUBERT [ 108
tragen. COFF1N (1928) stellte lest, dab die Singammer (Melospiza melodia) zum Tages- anbruch am besten und lebhaftesten (,,sprightly") singt, w~hrend spEterhin das Tempo langsamer und der Gesang leiser wird. ROBINSON (1949) sieht die D~immerungs- und Mondscheinges~inge australiseher SingvSgel als Anzeichen iiberm£Big starker Produktion yon Sexualhormonen an.
Der Fitislaubs~nger besitzt keine Gesangsthemen, die einer bestimmten Tages- zeit zugehSren, sondern es wurden nur Differenzen in der Gesangsl/inge, der H/iufigkeit von Wiederholungen, der Doppelstrophe und in dem prozentuellen Anteil einzelner Variationen entdeckt. Vergleichbare Themen variieren inter- individuell teilweise extrem unterschiedlich. D as effektorische Muster, welches diese Variationen produziert, kann yon einem M~innchen zum anderen in recht verschiedenartiger Korrelation zur endogenen Periodik und zur Gesangsaktivit/it stehen. J~hnliehe Themen mehrerer M~innchen k6nnen offenbar yon verschieden- artigen, circadian wechselnden Reizkonstellationen beeinfluBt werden. Nicht nur in der Gesangsaktivit~it, sondern auch in bezug auf zunehmende L~nge und die H~ufigkeitsverteilung des Doppelgesanges sind in der Mittagszeit Ten- denzen offensichtlich, welche ebenso in der sp~iteren Brutzeit auftreten. Morgens und abends herrscht dagegen mit Ausnahme des ,,Leierns" ein kurzer, pr~ignanter und intensiver Gesang vor. Die Wiederholungsrate korreliert im Tagesgang mit der GesangsaktivitEt teils positiv (5.--11. Stunde), teils deutlich negativ (11. bis 21. Stunde), jahreszeitlich jedoch positiv. Auch bei dem ,,Leiern" und einigen anderen Varianten stimmt das jahreszeitliche VerhEltnis zur Gesangsaktivit~it mit dem tageszeitlichen Muster nieht vollst~indig iiberein, so dab eine unmittelbare Korrelation von Aktivit~it und Auspr~igung der Strophe nicht in Frage kommen kann. Vielmehr handelt es sich im Jahresverlauf um endogen und exogen gesteu- erte Reifungsprozesse, die offenbar einen anderen Verlauf als die circadian ge- steuerten Vorg/inge, die der Motivwahl zugrunde liegen, einschlagen kSnnen. Ein weiterer Ankniipfungspunkt f/Jr diese Problematik wiirde sich aus der vergleichen- den Untersuchung der Lautst~rkeunterschiede im Tagesrhythmus und im Jahres- verlauf ergeben, die unserem Eindruck nach zeigen wfirde, daft bei groBer Akti- vit~it (morgens, abends und im April) lauter gesungen wird Ms bei niedriger (mittags und nach der Paarung) und dab somit der Wert des Gesanges als distanziiberbrfickendes Kommunikationsmittel und als den Revierbesitz anzei- gende Information in der Mittagszeit am geringsten ist.
I. Zusammenfassung Eine kleine Population von Fitislaubs~ngern (Phylloscopus trochilus [L.]) nSrdlich yon
Bernau bei Berlin wurde in der Brutzeit 1964 systematisch auf Gesangsvariationen und -aktivit~t untersucht. Besondere Beachtung fand die Korrelation der Gesangsaktivit~it zn den verschiedenen Variationen. Es konnte festgestellt werden, dab der Gesangsmodus durch Tageszeit, brutzeitliehen Status und die HShe der Singwarte beeinflul]t werden kann.
Benachbarte Individuen singen meist grundversehieden, und es konnte keine An- gleiehung der Ges~inge durch das gegenseitige Ansingen und HSren gefunden werden. Das Ansingen bedingt eine hohe und ausgeglichene H~ufigkeitsrate der Ges~inge der beteiligten Individuen.
Die t~igliche Beendigung des Gesanges streut naeh Helligkeit und Zeitpunkt st~irker als der Beginn. Die Phasenwinkeldifferenz A qo f/ir die Gesangsaktivit£t ist positiv. Sie nimmt

Heft 3] 1.967 ] Gesang des Fitis 289
in der Regal in den ersten Morgenstunden zu, fiillt vonder 8. bis zur 15. Stunde mit Unterbrechungen im Abstand yon jeweils 2 Stunden ab und erreieht am sp~iten Nachmittag wieder maximale Werte.
Die meisten Variationen sind im Verh~ltnis zum gesamten Repertoire tageszeitlich ver- schieden hiiufig. Positive Korrelation liegt besonders deutlieh bei den ,,Leierstrophen" eines M/innchens vor, negative Korrelation, welche jedoch in den Abendstunden in posi- tive umschl~gt, bei den Doppelstrophen. Die H~iufigkeit yon Wiederholungen verhKlt sich morgens in positiver Korrelation zur Gesangsaktivit~t, steigt dann von der 11. bis zur 16. Stunde an, f~illt in der 17. Stunde ab und steigt wiederum his zum Ende der Aktivi- tiitsphase an. Die Beziehung yon WiederholungshKufigkeit und Gesangsaktivit~it unter- liegt jedoch einer teilweise erheblichen individuellen Variationsbreite.
Der Gesang der friihen Morgan- und spiiten Abendstunden ist am kiirzesten. Der Wechsel von abgebrochenen und vielfachen Strophen im Revierkampf bewirkt eine sehr variable Gesangsliinge, die jedoch durch das Uberwiegen yon Abbriiehen kiirzer als in entspannten Situationen ist.
Mit der Besetzung des Reviers steigert Ph. trochilus die Gesangsratc und erreieht, wenn die Weibchen eintreffen, den HShepunkt. In der Balzzeit ist die Aktivitiit kurz- fristig gehemmt und wird nach einem darauffolgenden Anstieg friiher als bei unver- paarten M~innchen eingeschr'/inkt. Nach der Sommermauser ist wiederum ein kurzes aus- gepr/igtes Revierverhalten zu beobachten. Verpaarte Exemplare singen im allgemeinen variabler als ledige, deren Motive liinger als bei verpaarten beibehalten werden. Doppel- strophen erscheinen in der Regel in und nach der Paarungszeit besonders h~iufig und scheinen ein Ausdruck der epigamen Motivationskomponente zu sein, zumal da sie yon unverpaarten Hiihnen vial seltener gesungen werden. Sie treten allerdings auch im Revier- kampf auf, klingen dann jedoch anders und werden wahrscheinlich durch plStzliche Ent- hemmung der angestauten Singreaktion bewirkt. Das ,,Leiern", eine Verflachung der typischen Motive bei verpaarten Exemplaren, wurde in und nach der Balzzeit ebenfalls h~iufiger.
Mit fortschreitender Brutzeit wird die Strophenl/inge durch Phrasenverliingerung und den Einbau neuer Elemente sowie durch die zahlreichen Doppelges~inge grSller. W/ihrend fiir die Zeit der Revierbesetzung und -verteidigung ein relativ kurzer, lauter und priignanter Gesang typisch ist, wird er nach der Paarung liinger, leiser, weicher und lockerer. Die Strophigkeit wird abgebaut und die Kriterien des Subsong warden in zu- nehmendem MaBe deutlich.
Unter der Voraussetzung, dab der Gesang des Fitislanbs~ingers vornehmlich in der Zeit der Revierbesetzung und -verteidigung von arterhaltendem Wart ist, wird angenommen, dab die Ursache fiir die Tendenz zmn Subsong in der spiiten Fortpflanzungsphase in einem reduzierten Gestaltdruck liegt. Fernerhin unterliegt der Gestaltdruek einem tages- zeitlieh variierenden Muster.
S-mma.Ty
During the 1964 breeding season the song variations of a small population of Willow Warblers (Phylloscopus trochilus [L.]) in the north of Bernan near Berlin were in- vestigated systematically by means of tape recordings and auditory tests. The recordings were analysed by use of a sound spectrometer and a sonograph. In the auditory controls the song activity and the frequency of variations distinguishable by ear per 15 rain. were noted.
It was ascertained that the form of variations can be influenced by the time of day, the reproductive status and by the heigth of the singing perch. Particular attention was drawn to the correlation between song activity and the various modifications of the song:
The variations of neighbouring males are mostly very different from each other and there is no adjustment through countersinging and listening to each other. The mode of eountersinging affects a high, well-balanced level of song activity.

[J.Orn: 290 MICHAEL SCHUBERT [ 108
Average values were given to the brightness and the interval between sunrise and sunset, and the beginning and termination of the song activity respectively. The values disperse more in the end than in the beginning. The angle q~ of the phase of song activity is negative.
The level of the song activity as a rule rises in the early morning and declines from the eighth to the fifteenth hour of the day with intermittent breaks at intervals of two hours, developing thereafter a second maximum.
The percentage frequency of most variations changes in the course of the day. Positive correlation to the song activity could be found, for instance, in the "grinded" songs of a male, negative correlation in the double songs of all the investigated specimens, which changed to positive in the evening.
The frequency of repetitions of any song types in the morning corresponds to the song activity, rising from the tenth up to the sixteenth hour and, after a minimum, rising again; nevertheless, the relation varies from individual to individual.
The length of the song is shortest in the early morning and late evening. The preponderance of songs are broken off during territorial fights causing a certain shorten- ing. During the course of establishing the territory the song activity was at a maximum, culminating when the females arrive, with a short-term decrease during the mating time. Paired males sing thereafter Iess regularly than non-paired ones. After the post-nuptial moult a new marked rivalry is observable.
Paired males, in general, sing more variably than single ones, which retain some notes longer than the former.
Double songs are probably an expression of the epigamic component of motivation and the grinded songs occur conspicuously frequently during and after the courtship. Double songs, which occur in the aggressive comment, sound different from those resulting from a sudden re-activation of the congested singing reaction. The "grinded" songs, found only in non-paired males, become more frequent during and after the mating season.
The later the reproductive phase the longer the songs become through extension of the phrases (homologous elements), by insertion of new elements and by means of the double songs. In the days of establishment and defenee of the territory the mode of singing is relatively short, loud and terse whereas it becomes longer, lower and looser after the mating period. The context of a single song becomes reduced and characteristics of the subsong appear gradually.
If the song production of the Willow Warbler is especially essential during the time of occupation and defence of the territory, it is assumed that a diminished "Gestaltdruck" effects the subsong-tendency during the late reproductive period. Furthermore the "Ge- staltdruck" in the song varies in the course of the day being the strongest in the morning and evening.
L i t e r a t u r
ALLARD, N. (1930): The first morning song of some birds of Washington, D.C.: its relation to light. -- Amer. Nat. 64, p. 436--469.
- - (1934): The twilight song of the Wood Pewee, Myochanes virens Linnaeus. - Amer. Nat. 68, p. 266--274.
Am~rSTRONG, E. A. (1963): A study of bird song. London.
ASCrmFF, J. (1965): Circadian clocks. Amsterdam.
ASC~OFF, J., und R. W~.VER (1962): Beginn und Ende der t~iglichen Aktivitiit freilebcnder VSgel. -- J. Orn. 103, p. 2--27.
B~iKER, H. (1923): Der Gesang der VSgel und der periodische Ablauf der Spermiogenese. - J. Orn. 71, p. 169--196.

Heft 3] 1967 J Gesang des Fitis 291
BORROR, D. (1961): Intraspecific variation in Passerine bird songs. -- Wilson Bull. 73, p. 57--78.
BRAmCS, O. (1910): tiber einen bei Grevesmiihlen in Mecklenburg beobachteten Misch- s~inger (Phylloscopus trochilus rufus). -- Orn. Mschr. 35, p. 273--278.
BURGER, J. W. (1940): Further studies on the relation of the daily exposure to light to the sexual activation of the male Starling (Sturnus vulgaris). -- J. Exptl. Zool. 84, p. 351--360.
- - (1947): On the relation of day length to the phases of testicular involution and inactivity of the spermatogenetie cycle of the Starling. -- J. Exptl. Zool. 105, p. 259--267.
- - (19491: A review of experimental investigations on seasonal reproduction in birds. -- Wilson Bull. 61, p. 211--230.
- - (1953): The effect of photie and psychic stimuli on the reproductive cycle of the male Starling (Sturnus vulgaris). -- J. Exptl. Zool. 124, p. 227--239.
Buxro~, J. (1950): The Redstart. London.
C~RISTOLErr, E. (1907): Zur Frage des sogenannten Baumlaubvogels. -- Orn. Mschr. 32, p. 147--149.
COFFIn, L. V. B. (1928): Individuality in bird song. - Wilson Bull. 40, p. 95--99. CONDER, P.J. (1948): The breeding biology and behaviour of the continental Goldfinch
Carduelis c. carduelis. -- Ibis 90, p. 493--525. Cox, P. R. (1944): A statistical investigation into bird song. -- Brit. Birds 38, p. 3--9.
CRAIG, W. (1943): The song of the Wood Pewee Myiochanes virens Linnaeus: A study of bird music. -- N. Y. State Mus. Bull. 334, p. 1--186.
CUNNINGHAI~I, J.M. (1955): Diagrammatic expression of the seasonal intensity of bird song. -- Int. Orn. Congr. 11, p. 555--560.
CuP.re, E. (1959): Verhaltensstudien am Trauerschn~ipper. Berlin und Hamburg.
DAvis, J. (1958): Singing behavior and the gonad cycle of the Rufous-sided Towhee. - Condor 6, p. 308--336.
DAvis, J., und B. S. DAvis (1954): The annual gonad and thyroid cycles of the English Sparrow in Southern California. -- Condor 56, p. 328--345.
DELIUS, J. D. (1963): Das Verhalten der Feldlerche. - Z. f. Tierpsych. 20, p. 297--348.
DoP~o, C. (1924): Reizphysiologische Studien fiber den Gesang der V~igel im Hoch- gebirge. -- Pfliigers Arch. 204, p. 645-659.
FARKAS, T. (1955): Zur Brutbiologie und Ethologie des SteinrStels (Monticola saxatilis). - - Vogelwelt 76, p. 164--180.
FARNER, D.S. (1959): Photoperiodic control of annual gonadal cycles in birds. In: Photoperiodism and related phenomena in plants and animals. -- R. B. Withrow Ed. Publ. Am. Assoc. Adv. Science 55, p. 716--750.
FFR~Z, J. (1943): ~ber Ern~ihrung und Tagesrhythmus einiger VSgel im arktischen Winter. -- J. Orn. 91, p. 154--165.
- - (1949): Jahres- und Tagesrhythmus einiger Viigel in Nordfinnland. -- Z. f. Tierpsychol. 6, p. 308--329.
GRO~.~B~.LS, F. (1956): Untersuchungen iiber den morgendlichen Beginn und die abendliche Beendigung der Stimm~iul3erungen yon Vogelarten. -- Orn. Mitt. 8, p. 61--66.
HAARTm~, L. yon (1956): Territory in the Pied Flycatcher. -- Ibis 98, p. 460--475.
HAECK~,R, V. (1900): Der Gesang der VSgel, seine anatomischen und biologischen Grund- lagen. Jena.

I J .Orn 292 h~ICItAEL SCHUBERT [ 108
HAECKER, V. (1916): Reizphysiologisches tiber Vogelzug und Frfihgesang. - Biol. Zentral- blatt 36, p. 403--431.
- - (1924): Reizphysiologisches fiber den Abendgesang der VSgel. -- Pfliigers Arch. 204, p. 718--725.
HARTSnORNE, C. (1956): The monotony threshold in singing birds. -- Auk 73, p. 176--192. HOESCH, W., und G. NIETHAMlYIER (1940): Die Vogelwelt Deutsch-Siidwestafrikas, nament-
lich des Damara- und Kamalands. -- J. 0rn. 88, Sonderheft, p. 1--104.
HOF~ANN, K. (1959): l_lber den Tagesrhythmus der VSgel im arktischen Sommer. - J. Orn. 100, p. 84 -89 .
HOMANN, P. (1960): Beitr~ige zur Verhaltensbiologie des Weidenlaubsiingers (Phyllos- copus coUybita). -- J. Orn. 101, p. 195--224.
HOMA~N, P., und E. GWINNER (1963): Zum Balzverhalten des Zilpzalps, Phylloscopus collybita, im Friihling und im Herbst. -- J. Orn. 104, p. 365--371.
KAISER, W. (1965): Der Gesang der Goldammer und die Verbreitung ihrer Dialekte. - Der Falke 12, p. 40--42, 92--93, 131--135, 169--170.
KALELA, 0. (1958): (~ber aul3erbrutzeitliches Territoria]verhalten bet VSgeln. - Ann. Acad. Scient. Fennica, Ser. A, IV Biolog. H. 42.
KLOCKARS, B. (1941): Studier 5ver f~gelsfingens dagsrhythmik. -- Ornis Fennica 18, p. 73--110.
KuvslSTO, P. (1941): Studien fiber die (~kologie und den Tagesrhythmus von Phyllos- copus trochilus acredula (L.). -- Acta Zool. Fennica 31, p. 1--120.
LANYON, W . E . (1957): The comperative biology of the Meadowlarks (Sturnella) in Wisconsin. -- Pub]. Nuttall. Orn. Club 1, p. 1--67.
LEHVONEN, L. (1954): The song rhythm and calls of the Great Tit at various times of the year. -- Orn. Fennica 31, p. 99--115.
LENNERSTEDT, I. (1964): Nfigra drag i h~ickningsbiologin hos 15vs~lngare, busksviitta och s~ivsparv i mellersta Lappland. -- Fauna och Flora 59, p. 94--123.
L~OPOLD, A., und A. E. EYNON (1961): Avian daybreak and evening song in relation to light intensity. -- Condor 63, p. 269--293.
MARLER, P., und D. ISAAC (1960): Physical analysis of a simple bird song exemplified by the Chipping Sparrow. -- Condor 62, p. 124-135.
MARSHALL, A . J . (1951): The refractory period of the testis rhythm in birds and its possible bearing on breeding and migration. -- Wilson Bull. 63, p. 238--261.
- - (1952): The interstitial cycle in relation to autumn and winter sexual behaviour in birds. -- Proc. Zool. Soc. London 121, p. 727--740.
- - (1960): The environment, cyclical reproductive activity and behaviour in birds. -- Symp. Zool. Soc. LoNdon 2, p. 53--67.
MARSHALL, F. A. H. (1936): Sexual periodicity and the causes which determine it. -- Phil. Trans. Roy. Soc. London. Series B, 266, p. 423--456.
MAY, D . J . (1949): Studies on a community of Willow Warblers. -- Ibis 91, p. 24--54.
M~t~KSL, F. (1956): Untersuchungen fiber tages- und jahresperiodische Aktivit~itsiinderun- gen bet gck~figten Zug~Sgeln. -- Z. f. Tierpsychol. 13, p. 278-301 .
- - (1958): Untersuchungen fiber tagcs- und jahresperiodische ~nderungen im Energiehaushalt gekiifigter ZugvSgel. -- Ztschr. vergl. Phys. 41, p. 154--178.
MEssYmn, E., und I. MESSMER (1956): Die Entwicklung der Lant~ul3erungen und einiger Verhaltensweisen der Amscl (Turdus merula merula L.) unter natiirlichen Be- dingungen und nach Einzelaufzucht in schalldichten R~iumen. - Z. f. Tier- psychol. 13, p. 341--441.

Heft 3] 1967 ]
NIcs, M.
PALM, B.
Gesang des Fitis 293
(1931): Notes on twilight songs of the Scissor-tailed and Crested Flycatcher. -- Auk 48, p. 123--125. (1943): Studies in the life history of the Song Sparrow. II. The behaviour of the Song Sparrow and other passerines. -- Trans. Linn. Soe. N. Y. 6, p. 1--328.
(1951): Efter~rssangcn hos Gransanger (Phylloscopus collybita [Vieill.]). -- Dansk. orn. For. Tidskr. 45, p. 139--144.
PALMGREN, P. (1932): Ein Versu& zur Registricrung der IntensitStsvariation des Vogel- sanges im Laufe eines Tages. -- Orn. Fenniea 9, p. 68--74.
- - (1934): Balz als Ausdruek der Zugextase bei einem gek~ifigten Fitislaubs~inger. - - Orn. Fennica 11, p. 84--87.
POULSBN, H. (1958): The calls of the Chaffinch (Fringilla coelebs L.) in Denmark. -- Dansk. Orn. For. Tidskr. 52, p. 89--105.
ROBINSON, A. (1949): The biological significance of bird song in Australia. -- Emu 48, p. 291--315.
ROWAN, M.K. (1955): The breeding biology and bchavio~r of the Red-winged Starling Onychognathus morio. -- Ibis 97, p. 663--705.
SAUEa, F. (1955a): 1Jber die Variationen der Artges~inge bei Grasmiieken. -- J. Orn. 96, p. 129--146.
- - (1955b): Zmn Problem der Grasmiiekenges~inge. -- Orn. Mitt. 7, p. 161--165.
SAVNDSRS, A .A. (1947): The seasons of bird song. The beginning of song in spring. -- Auk 64, p. 97--I07.
- - (1948a): Theseason of bird song -- the cessation of song after the nesting s e a s o n . - Auk 65, p. 19--30.
- - (1948b): The seasons of bird song. Revival of song after the postnuptial molt. -- Auk 65, p. 373-383 .
SCHEER, G. (1941): iJber den Frtihgesang der VSgel. -- Verh. Orn. Ges. Baycrn 22, p. 137--159.
SCHmD¢, G. (1957): Abweiehende Laut~iul~erungen des Fitis. -- Vogelwelt 78, p. 37--38.
SCHiiZ, E. (1950): Am Brutplatz des Griinen Laubs~ingers (Phylloscopus trochiloides viridanus Blyth.). -- In: Syllegomena biologica. Leipzig, Wittenberg.
SCHWAN, A. (1920): Vogelsang und Wetter, physikaliseh-biologisch u n t e r s u e h t . - Pfliigers Arch. 180, p. 341--347.
- - (1921--23): t iber die Abh~ingigkeit des Vogelgesanges yon meteorologisehen Fa!ctoren, untersueht auf Grund physikaliseher Methoden. -- Verh. Orn. Ges. Bayern 15, p. 9--42, 146--186.
STIEVE, H. (1950): Der Gesang der VSgel und seine Abh/ingigkeit yon den Ke imdr i i s en . - Syllegomena biologiea. Leipzig, Wittenberg.
TEM~OCI¢, G. (1963): Mischlaute beim Rotfuehs (Vulpes vulpes L.). -- Z. f. Tierpsyehol. 20, p. 616--623.
THn~LCKE, G., und K. E. LI~S~NMAm (1963): Zur geographischen Variation des Gesanges des ZilpzMps, Phylloscopus collybita, in Mittel- und Siidwesteuropa. mit einem Vergleich des Gesanges des Fitis, Phylloscopus trochilus. -- J. Orn. 104, p. 372--402.
THORPE, W . H . (1958): The learning of song patterns by birds, with especial reference to the song of the Chaffinch Fringilla coeIebs. -- Ibis 100, p. 535--570.
VLm~, L. (1961): Observations on the daily rhythm of the song of some forest birds in central and northern Sweden. -- Ardea 49, p. 158--164.

[J.Orn. 294 MICHAEL SCHUBERT [ 108
WAGNER, H.O. (1956): Uber Jahres- und Tagesrhythmus bei ZugvSgeln. -- Z. f. Tier- psychol. 13, p. 82--92.
WOLFSON, A. (1941): Light versus activity in the regulation of the sexual cycles of birds.: The rSle of the hypothalamus. -- Condor 43, p. 125--136.
- - (1952): Day length, migration and breeding cycles in birds. -- Sci. Mon. N. Y. 74.4, p. 191-200.
WmGHT, H. W. (1913): Morning awakening and evening-song. -- Auk 30, p. 512--537.
ZIMMER, C. (1920): Der Beginn des Vogelgesanges in der Frfihdiimmerung. - Verh. Orm Ges. Bayern 14, p. 152--180.





























