Embed Size (px)
Citation preview

BIOCHIM1E, 1973, 55, 5-10. M6moires originaux
Glycosidases de Dolichos biflorus. Isolement et propri6t6s g6n6rales.
Denise MEYER (*) et Roland BOURRILLON.
L a b o r a t o i r e de B i o c h i m i e - - C e n t r e de R e c h e r c h e s s u r les P r o t d i n e s (**), F aca l t ~ de M ~ d e c i n e , ~5, rue des Sa in l s -P~res , P a r i s 6 ~, F r a n c e .
( 1 5 / 5 / 1 9 7 2 ) .
Summarl l . - - The purification of the osidases, ~-N-aeetylglueosaminidase, ~- and [~-galactosidases, ct-mannosidase and ~-glucosidase from the germinating seeds of Dolichos bi f lorus is described. These enzymes have been isolated in a highly active form.
The purification involved extraction, ammonium sulfate fraetionation and chromato- graphy on DEAE-Sephadex A-50.
The enzyme preparations were unstable to freezing and thawing but only t~-mannosi- dase activity was lost by lyophilisation ; all the enzyme preparations but c~-galactosidase and especially ~-glucosidase were stable to storage at 4 °. The pH optimum of each glyeo- sidase has been determined with o- or p-nitrophenyl D-glucopyranosides as substrates.
INTRODUCTION.
Les gIycosidases sont largement distr ibu6es chez les animaux, les v6g6taux et les microorga- nismes [1-10J. De nombreux t ravaux ont 6t6 con- sacr6s aux osidases des tissus mammif6res [1-3] et, r6cemment, l ' a t tent ion a 6t6 attir6e par les osidases v6g6tales [7=10~. En effet les extrai ts aqueux des graines de P h a s ~ o l u s mdf far i s [10! r6v616rent des activit6s ~-glucosidase, a -mannos i - dase, [~-N-ac6tyl-hexosaminidase, a- et ~-galactosi- dases tr6s 61ev6es. En outre, ces osidases aug- menten t net tement au cours de la germinat ion qui se caract6rise par la d6gradat ion de biopo- lymbres polysacchar idiques , ce qui fourni t l '6ner- gie et les in te rm6dia i res pour le d6but de la crois- sance embryonna i re .
L ' int6r6t de la purif icat ion, de la caract6ri- sation et de la sp6cificit6 des osidases est li6 h leur usage possible dans la d~terminat ion de la s6quence et de la configurat ion anom6rique des rdsidus carbohydra tes dans les complexes hdt6- rosacchar id iques [11].
L 'examen d 'un extrait b ru t aqueux des graines de D o l i c h o s b i f l o r u s montre l 'exis tence d 'une forte activit6 de la ~-galactasidase (E.C.3.2.1.23), de la ~-N-ac6tyl-glucosaminidase (E.C.3.2.1.30) et de l 'a- mannos idase (E.C.3.2.1.24) et, /I un degr6 moindre , de l 'a-galactosidase (E.C.3.2.1.22) ainsi que des a- et ~-glucosidases (E.C.3.2.1.20, E.C.3.2.1.21).
(*) Charg6e de Reeherehes au CNRS. (**) Equipe de Reeherches assoeide au Centre Na-
tional de la Recherche Scientifique.
Le pr6sent t ravai l rappor te la pur i f icat ion et quelques propri6t6s de c inq osidases extraites des graines de D o l i c h o s b i f l o rus , phytoh6maggluti- n ine h activit6 sp6cifique (A1) de groupe sanguin, dont les activit6s osidasiques augmentent consi- d6rablement au cours de la germinat ion.
MATERIEL ET METHODES.
Mater ie l .
Les graines de D o l i c h o s b i f l o r u s prov iennen t de Ahmedabad (Inde).
Le DEAE-S6phadex A-50 est obtenu chez Phar- macia, Uppsala (Su6de).
Les substrats enzymat iques : o-nitroph6nyl-g-D- glucopyranoside, p -n i t roph6ny l -a -D-ga lac topyra - noside, p-ni troph6nyl<t- et ~-D-glucopyranosides, p -n i t roph6nyl -a -D-mannopyranos ide et p-ni troph6- nyl-2-ac6tamido-2-d6oxy-g-D-glucopyranoside sont fournis par Koch-Light - - Laborator ies LTD.
M ~ t h o d e s .
- - G e r m i n a t i o n : Les graines s~ches sont lav6es duran t 15 minu tes par agitat ion dans une solution de HgCL, h 0,1 p. cent, puts r inc6es h l 'eau distill6e et enfin mises h gonfler pendan t 24 heures dans l 'eau distill6e [12].
Aprbs s6chage rapide, les graines sont raises /t germer dans des plateaux entre des feuilles de papier Wha tman N ° 3 humidifi6es. Les plateaux, recouverts d 'une plaque de verre, sont ma in tenus • h l 'obscuri t6 et h temp6rature ambiante pendan t p lus ieurs jours.
6 D e n i s e M e y e r e t R o l a n d B o u r r i l l o n .
- - D E A E - S ~ p h a d e x A-50 : Le DEAE-S6phadex A-50 est mis h gonfler dans l 'eau distill6e puis mis en suspension dans le tampon phosphate de so- d ium 0,04 M h pH 6,5. Une colonne de ce DEAE- S6phadex A-50 (2 ,5 )< 70 cm) est 6quilibr6e -t- 4°C, dans le m6me tampon phosphate de sodium.
Aprbs d6p6t de l '6chant i l lon, l'61ution est r6ali- s6e h l 'a ide d 'un gradient 6tabli au moyen d 'un varigrad, entre 600 ml de tampon phosphate de sodium 0,04 M h pH 7,5 et 600 ml de m6me tampon r e n d u 1 M e n NaC1.
- - Dosages d e s p r o t d i n e s : Les prot6ines sont dos6es soit pa r la m6thode du b iure t [13], soit par la m6thode de Lowry et al. [14], avec la s6rum a lbumine bovine comme s tandard.
- - Dosages e n z y m a t i q u e s : Les activit6s enzy- matiques sont estim6es selon le proc6d6 de Conchie et Hay I15] ; chaque enzyme est test6e h son pH op t imum dans le t ampon Na2HPO ~ 0,2 M-acide ci t r ique 0,1 M ( tampon de McIlvaine) I~63.
0,15 ml de la solution d 'enzymes ~ une di lut ion convenable sont ajout~s au m~lange des solutions de tampon et de substrat ad~quates, le volume final incub6 ~tant de 1,2 ml. La concent ra t ion finale en substrat dans le mil ieu incub~ est de 2,5 mM pour l 'o-ni t rophdnyl-~-D-galactopyrano- side, 6 mM pour le p-n i t roph~nyl -a-D-mannopy- ranos ide et 5 mM pour les autres p-ni t roph~nyl- glycosides.
Le m~lange est ineub~ au ba in-mar ie h 37°C pendan t 1 heure, et la r~action est arrSt6e par addi t ion de 1,2 ml de t ampon glycine-NaOH 0,4 M pH 10,8. La quanti t6 de p- ou o-ni t roph~nol lib~r6 est d~termin~e h 430 n m pa r l ' intensi t~ de la colo- ra t ion jaune form~e, par rappor t h une courbe de r6f~rence ~tablie de fa~on s imilaire avec des prises d'essai de 0,09 h 0,3 ~M d'o- ou p-ni t ro- phenol.
Les pH opt ima ehoisis pour les dosages sont 3,9 pour la ~-galaetosidase et l 'u-mannosidase , 4,5 pour les a- et ~-glucosidases, 4,6 pour la ~-N- ac~tylglucosaminidase et 5,1 pour l 'a-galactosi- dase.
- - D ~ f i n i t i o n de s un i t~s : L 'uni t~ d 'enzyme est d~finie comme la quantit~ qui lib~re 1 .uM d'o- ou p-nitroph~,nol par minu te h 37°C. L'activit~ sp~ci- fique de l ' enzyme est exprim~e en unit~ d 'enzyme par mg de prot~ine.
RI~SULTATS.
I - - P u r i f i c a t i o n de s e n z y m e s .
Toutes les opera t ions d 'ext rac t ion et de purifi- cat ion sont effectu~es h q- 4°C.
E x t r a c t i o n .
50 g de graines s~ches sont mis /i germer pendan t 6 jours fi 16°C. Les gra ines germ~es sont f inement broy~es, puis homog~n~is~es dans 2,5 fois leur poids de NaC1 fi 0,9 p. cent. La suspension est agit~e pe nda n t 24 heures, filtr~e sur gaze et centrifug~e dans une centr i fugeuse Sorvall Rc-2B r~frig~r~e, h 9 500 × g pe nda n t 1 heure. Le l iquide clair surnageant (470 ml) est recueil l i .
F r a c t i o n n e m e n t p a r le s u l f a t e d ' a m m o n i u m .
L'extrai t aqueux de l 'op6rat ion pr~c~dente est addi t ionn~ de sulfate d ' a m m o n i u m cristallis~ jusqu'h la molarit~ 3,2. Le pr~cipit~, recuei l l i par centr i fugat ion, est repr is dans 130 ml de tampon phosphate de sodium 0,2 M h pH 6,5. Un l~ger insoluble est ~cart~ par centr i fugat ion. Le surna- geant est repr~cipit~ par le sulfate d ' ammonium, dans les m~mes c o n d i t i o n s ; le pr~cipit~ form~ est redissout dans 120 ml de tampon phosphate
T x ~
20 ~O eo 8 0 I o0 120 ~40 leO l e o
FRACTION ( NUMIERO )
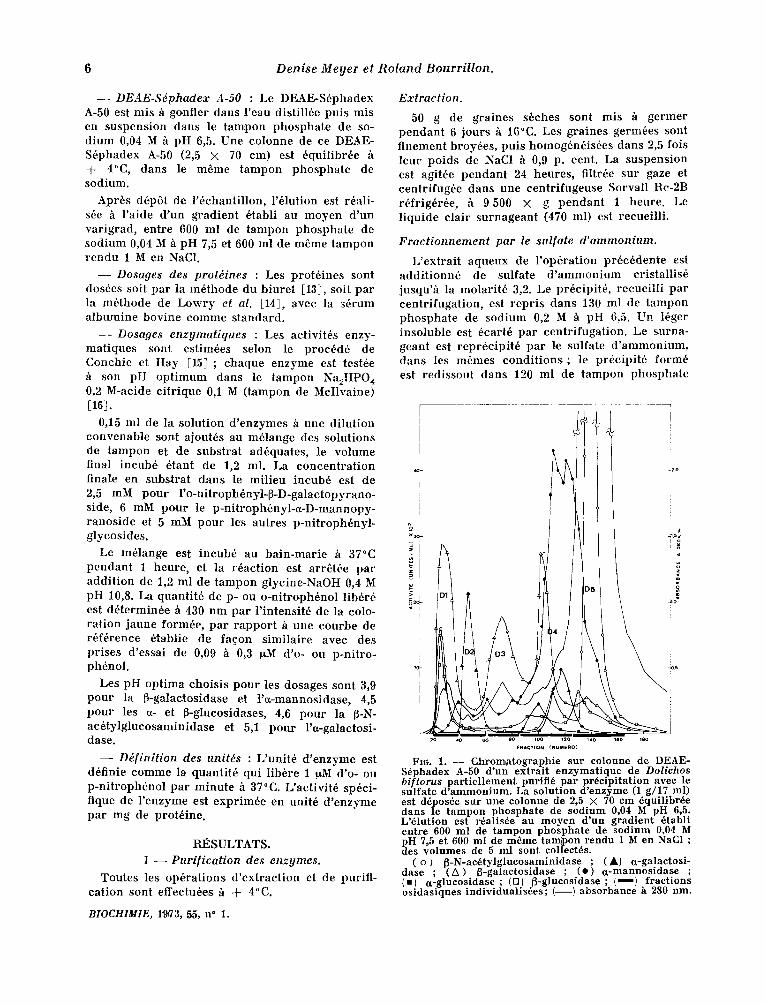
FIG. 1. - - Chromatographic sur coloune de DEAE- S~phadex A-50 d'un extrait enzymatique de Dolichos bi t lorus partiellement purifi~ par precipitation avec le sulfate d'ammonium. La solution d'enzyme (1 g/17 ml) est d~pos~e sur une c o l o n n e de 2,5 × 70 cm ~qui l ibr~e dans le tampon phosphate de sodium 0,04 M pH 6,5. L'~lut ion es t r~al i s~e au m o y e n d'un g r a d i e n t ~tabl l entre 600 ml de tampon phosphate de sodium 0,04 M pH 7,5 et 600 ml de m~me tam~pon rendu 1 Men NaC1 ; des volumes de 5 ml sont collect~s.
( o ) ~ - N - a c ~ t y l g l u c o s a m i n i d a s e ; ( A ) u -ga l ac to s i - dase ; C A) ~-galactosidase ; Co) ~-mannosidase ; (m) ~ - g l u e o s i d a s e ; (D) ~ - g l u c o s i d a s e ; ( ~ ) fractions o s i d a s i q u e s i n d i v i d u a l i s ~ e s ; ( - - ) a b s o r b a n e e h 280 nm.
BIOCHIMIE, 19~73, 55, n ° 1.
Glycosidases de D o l i c h o s b i f lo rus .
de sodium 0,04 M h pH 6,5 ; la solution obtenue est amen6e h la molari t6 4,0 en sulfate d ' ammo- n ium et le nouveau pr6eipi t6 est trait6 eomme ei- dessus. F ina lement un extrai t enzymat ique de volume 164 ml (D-P 80) est obtenu, sur lequel les diverses aetivit6s osidasiques seront d6termin6es.
Concentration par dialyse sous vide.
L'ext ra i t enzymat ique D-P 80 est concentr6 h faible volume par dialyse sous vide (tube h dialyse - - Visking 8/32) contre le tampon phosphate de sodium 0,04 M pH 6,5.
Chromatographie sur DEAE-S~phadex A-50.
Apr6s centr i fugat ion, la solution enzymat ique concentr6e (1 g/17 ml) est d6pos6e sur la colonne de DEAE-S6phadex A-50.
Apr6s 6tablissement du gradient d'61ution, des f ract ions de 5 ml sont recueil l ies, h la vitesse de 10 ml /heu re . Pour un tube sur deux la concen- t ra t ion en prot6ines (absorpt ion U.V. h 280 nm) et les aet ivi t6s enzymat iques sont d6termin6es. La figure 1 repr6sente le profil d'61ution obtenu :
sidase et de l 'a-glueosidase, ainsi que de faibles aetivit6s ~-galactosidase III, ct-galactosidase II, fi-glucosidase II et .~-N-ac6tylglucosaminidase.
- - La f ract ion D5 (tubes 196 "~ 165), f i che en prot6ines, se caract6r ise surtout par la pr6senee de fi-N-ac6tylglucosaminidase, encore contamin6e fa ib lement pa r l 'ct-mannosidase et la ~-glucosi- dase III.
Ces diff6rentes f ract ions sonl concentr6es par dialyse sous vide et conserv6es h 20"C h l'6tat eongel6. I1 a 6t6 constat6 que cer ta ines activit6s osidasiques, en par t i eu l i e r a-mannosidasiques , 6taient d6truites par lyophi l isa t ion.
I1 est observ6 que cer ta ines aetivit6s osidasiques se re t rouvent dans deux ou trois pies diff6rents. C'est le eas no tamment de la ~-galaetosidase pr6- senle en D1, D3 et D4, de l'¢t-glueosidase (D2 et D4) et de la ~-glucosidase (D1, D4 et DS).
Les rendements , expr im6s en activit6 enzyma- tique, sont g6n6ralement eompr i s entre 60 et 85 p. cent, sauf dans le eas des ct- et ~-galaetosidases dont les rendements sont respee t ivement 34 et 46 p. cent.
T A B L E A U I .
Purification de glycosidases de Dolichos biflorus.
Extrait brut . . . . Prdeipit~ sulfate Dialyse sous vide
DEAE-Sx A-50
~.-6alaetosidase [ ~-6alaelosidase
Aetiviti tatale
unitds
87 83 58
I II
1 4 6
Aetivit6 Aetivit6 Aetivit6 ~p6eifique totale sptieifique Iotale
unitt%/mg [ unit6s unitds ling [ unitds x 103 ] x 10 :~ !
33 I 159 59 i 35
57 I1 I 149 51 140
[ I I I l l
46---9 4"5 ~ 17 21
102 ] 27
122 II I 24 1 I I I
11128 370 ~
~-Glueosidase I
ietivil~ Aetivit6 sp6cifique
unilds/mg x 10 ~
13 1~ 21
I
406
7-Mannosidase
A¢litil~ 101Me
unit6s
115 111 111
pic majeur
4;s
~-N-Ae6tylgluco- saminidase
Aetivit6 Atlifili Aetivil~ speeifique 10tale ~p~eifique
tmitds/mg i unit6s unitds/mg x 10 :~ x 10:2
43 140 52 75 138 94 97 139 122
pic majeur
85 ! 132
Les chiffres sont rapport~s h 50 g de graines s6ehes, mises h germer pendant 6 jours.
- - La f rac t ion D1 (tubes 20-39) se earact6r ise par la pr6sence d 'une ~-galactosidase I avee des activit6s ~-glueosidase I e t ~-N-ae6tylglucosamini- dase net tement plus faibles.
- - L a f rac t ion D2 (tubes 41 h 60) cont ien t presque exc lus ivement une act ivi t6 a-galaetosi- dase I.
- - L a fract ion D3 (tubes 62 h 85) prbsente essent ie l lement une aetivit6 ~-galaetosidase II.
- - La f rac t ion D4 (tubes 86 h 125) est h6t6ro- g6ne. Elle eont ient la quasi-totalit6 de l '~-manno-
Le tableau I r appor te les activit6s totales et sp6- cifiques des diff6rentes osidases r6solues par ehro- matographie sur DEAE-S6phadex A-50.
II - - Propri~t~s des osidases.
Germination,
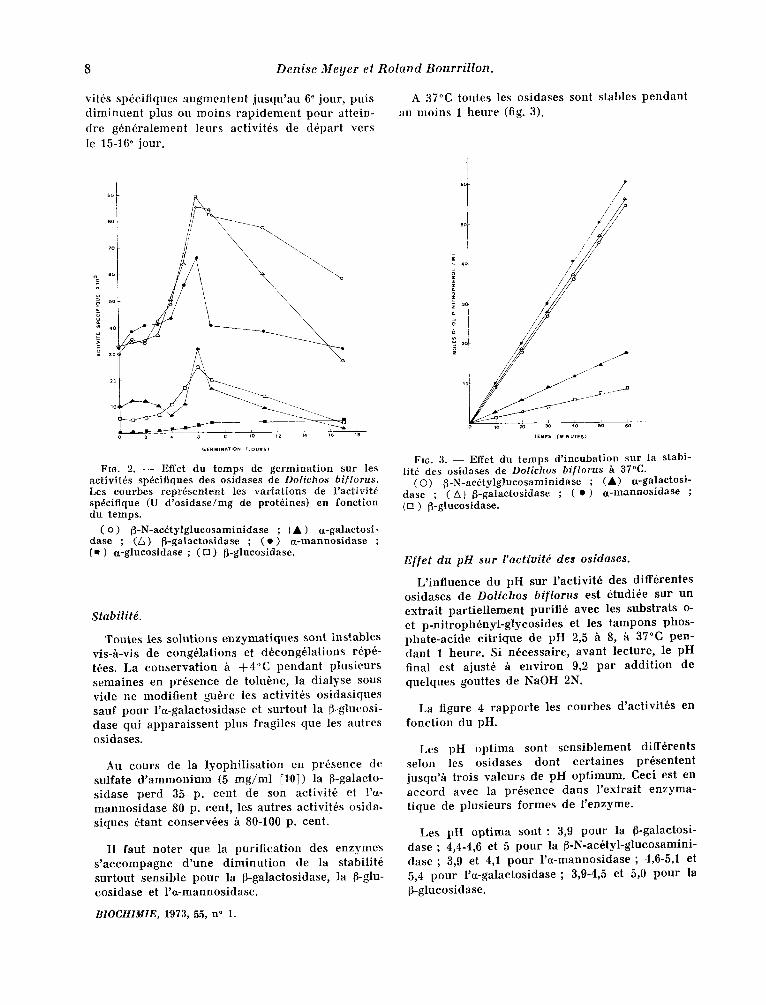
La figure 2 rappor te l ' inf luenee du temps de germina t ion sur les aetivit6s enzymat iques sp6ei- t iques des diverses o s idase s ; ees essais sont r6alis6s /a pa r t i r de divers extrai ts bruts dialys6s contre l 'eau distill6e. I1 est observ6 que les aeti-
BIOCHIM1E, 19q3, 55, n ° 1.
Denise Meyer et Roland Bourri l lon.
vitds spdcifiques augmentent jusqu 'au 6 ° jour, puis d iminuen t plus ou moins rap idement pour attein- dre gdndralement leurs activitds de ddpart vers le 15-16 ° jour.
I
° i 7O
~ER M~NATION [JOURS~
Fro. 2. - - Effet du temps de germination sur les aetivit~s spdeifiques des osidases de Dolichos biflorus. Les eourbes repr~sentent les variations de Yactivit~ spdcifique (U d'osidase/mg de protdines) en fonetion du temps.
( o ) fJ-N-aedtylglueosaminidase ; [ • ) a-galaetosi- dase ; (/,) ~-galaetosidase ; ( e ) a-mannosidase ; ( • ) a-glucosidase ; ( [] ) [}-glucosidase.
Stabilit&
Toutes los solutions enzymat iques sont instables vis-A-vis de congdlations et ddcongdlations rdpd- tdes. La conservat ion A + 4 ° C pendan t p lus ieurs semaines en prdsence de toluene, la dialyse sous vide ne modifient gu~re los activitds osidasiques saul pour l 'a-galactosidase et surtout la ~-glucosi- dase qui apparaissent plus fragiles que los autres osidases.
Au cours de la lyophi l i sa t ion en prdsence de sulfate d ' a m m o n i u m (5 mg /ml [10]) la ~-galacto- sidase perd 35 p. cent de son activitd et l'u- mannos idase 80 p. cent, les autres activitds osida- siques dtant conserv~es h 80-100 p. cent.
I1 faut noter que la pur i f icat ion des enzymes s 'accompagne d 'une d iminu t ion de la stabilitd surtout sensible pour la ~-galactosidase, la I}-glu- cosidase et l 'ct-mannosidase.
A 37°C toutes les osidases sont stables pendan t au moins 1 heure (fig. 3).
5
t /
O
6
o o 70
TEMPS {MINUTE$}
Fro. 3. Effet du temps d'incubation sur la stabi- litd des osidases de Dolichos billorus h 37°C.
(O) ~-N-aedtylglncosaminidase ; (A) a-galaetosi- dase ; (A} ~-galaetosidase ; ( • ) a-mannosidase ; (0) ~-glueosidase.
E[[et du pH sur l'activit~ des osidases.
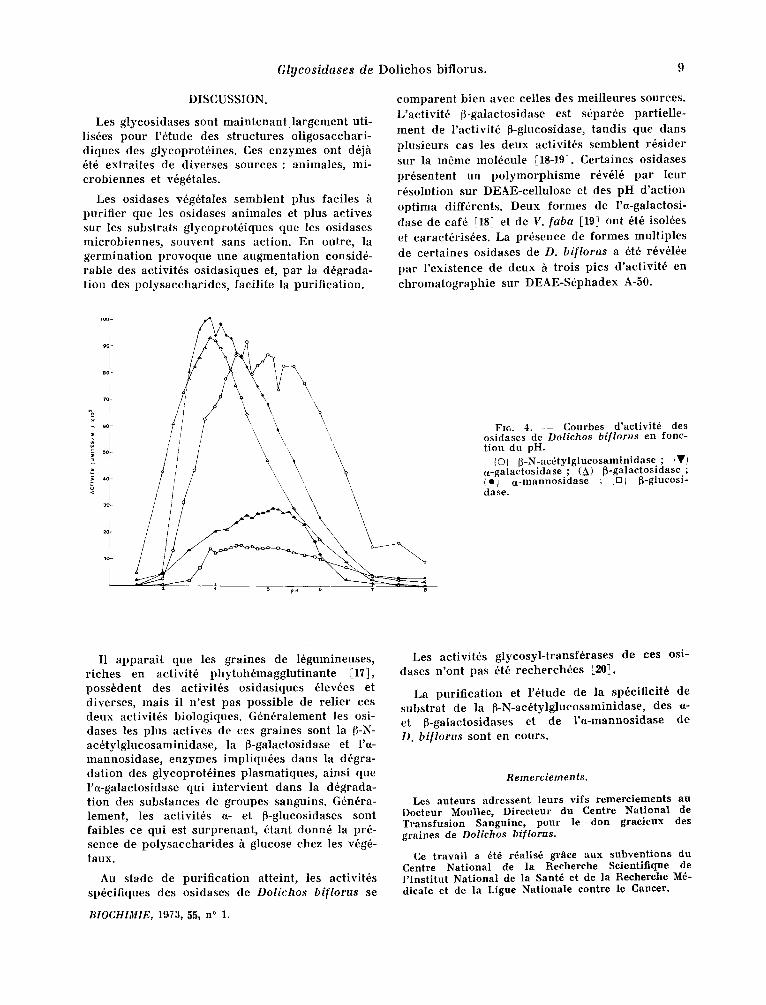
L'inf luence du pH sur l 'act ivi td des diff~rentes osidases de Dolichos bi[lorus est dtudide sur un extrai t par t ie l lement purifid avec los substrats o- et p-ni t rophdnyl-glycosides et los tampons phos- phate-acide ci t r ique de pH 2,5 ~t 8, ~ 37°C pen- dant 1 heure. Si ndcessaire, avant lecture, le pH final est ajustd h envi ron 9,2 par addi t ion de quelques gouttes de NaOH 2N.
La figure 4 rappor te los courbes d'activit~s en fonct ion du pH.
Los pH optima sont sensiblement diffdrents solon les osidases dont certaines pr~sentent jusqu'h trois valeurs de pH opt imum. Ceci est en accord avec ]a prdsence dans l 'extrai t enzyma- tique de plusieurs formes de l 'enzyme.
Les pH opt ima s o n t : 3,9 pour la ~-galactosi- dase ; 4,4-4,6 et 5 pour la ~-N-acdtyl-glucosamini- d a s e ; 3,9 et 4,1 pour l ' a -mannosidase ; 4,6-5,1 et 5,4 pour l 'a-galactosidase ; 3,9-4,5 et 5,0 pour la ~-glucosidase.
BIOCHIMIE, 1,973, 55, n ° 1.
Glycosidases de Dol ichos b i f lorus .
DISCUSSION.
Les glycosidases sont ma in t enan t largement uti- lis6es pour l '6tude des s t ructures oligosacchari- diques des glycoprot6ines. Ces enzymes ont d6j'a 6t6 extrailes de diverses sou rce s : animales, mi- c robiennes et v6g6tales.
Les osidases v6g6tales semblent plus faciles pnr i f ier que les osidases animales et plus a t t i r e s sur les substrats glycoprot6iques que les osidases microbiennes , souvent sans action. En ou_fre, la germinat ion provoque une augmenta t ion consid6- rable des activit6s osidasiques et, par la d6grada- t ion des polysaccharides, facilite la purif icat ion.
comparent bien avec celles des meil leures sources. L'activit6 ~-galactosidase est s6par6e partielle- ment de l 'activit6 g-glucosidase, landis que dans plusieurs cas les deux activit6s semblent rSsider sur la m6me nml6cule [18-19]. Certaines osidases pr6sentent un poly inorphisme r6v616 par leur r6solution sur DEAE-cellulose et des pH d 'act ion optima diff6rents. Deux fonnes de l 'a-galactosi- dase de car6 [18] et de V. [aba [19] ont 6t6 isol6es et caract6ris6es. La pr6sence de formes mult iples de certaines osidases de D. biflorus a 6t6 r6v616e par l 'existence de deux ~ trois pics d 'act ivi t6 en chromatographie sur DEAE-S6phadex A-50.
i oo
gc
7O
SO
/,
/
\ \ \
FIr . 4. - - Courbes d'activit~, des os idases de Dolichos biflorus cn fonc- tion du pH.
(O~ ~-N-ac6tylglucosaminidasc ; IWI ct-galactosidase ; (A) 6-galactosidase ; (e~ a-mannosidase : ~D/ ~-glucosi- dasc.
I1 appara i t que les graines de 16gumineuses, r iches en activit6 phytoh6magglu t inante [17], poss~dent des activit6s osidasiques 61ev6es et diverses, mats il n 'est pas possible de rel ier ces deux activit6s biologiques. G6n6ralement les osi- dases les plus actives de ces graines sont la g-N- ac6tylglucosaminidase, la ~-galactosidase et l 'a- mannosidase , enzymes impliqu6es dans la d6gra- dat ion des glycoprot6ines plasmatiques, a insi que l 'a-galactosidase qui in te rv ien t darts la d6grada- l ion des substances de groupes sanguins. G6n6ra- lement, les activit~s a- et g-glucosidases sont faibles ce qui est su rp renan t , 6tant donn6 la pr6- sence de polysacchar ides fi glucose chez les v6g6- taux.
Au stade de puri f icat ion atteint, les activit6s sp6cifiques des osidases de Dolichos biflorus se
BIOCHIMIE, 1973, 55, n ° 1.
Les activit6s glycosyl-transf6rases de ces osi- dases n 'on t pas 6t6 reeherch6es [20].
La puri f icat ion et l '6tude de la sp6cificit6 de substrat de la ~-N-ac6tylglucosaminidase, des a- et ~-galactosidases et de l ' a -mannosidase de D. bi[Iorus sont en cours.
Remerciements.
Les a u t e u r s ad re s sen t l eurs v i fs r e m e r c i e m e n t s an Doc teur Moullec, Di rec teur du Centre Na t iona l de T r a n s f u s i o n Sanguine , p o u r lc don grac icux des gra ines de Dolichos biflorus.
Ce t rava i l a 6td r6alis6 grace aux s u b v e n t i o n s du Centre Na t iona l de la Recherche Scientif lque de l ' I n s t i t u t Na t iona l de la Sant~ et de la Recherche M~- dicale ct de la Ligue Nat iona l¢ contre le Cancer,

10 D e n i s e M e y e r et R o l a n d B o u r r i l l o n .
R~SUM~.
La purification des osidases ~-N-ac~tylglucosamini- dase, a- et ~-galactosidases, a -mannos idase et ~-glu- cosidase, ~t par t i r de graines germ4es de Dotichos bi- [lorus, est d~crite. Ces enzymes ont 6t~ isol~es sons une forme tr~s active.
La purification se fai t eu trois ~tapes : extraction, f rac t ionnement par le sulfate d ' ammonium et ehroma- tographie sur DEAE-S4phadex A-50.
Les preparat ions enzymatiques sont instabtes vis-h- vis de cong~lations et ddcongdlations suecessives, mais seule l 'activit4 u-mannosidas ique est perdue par lyophil isat ion ; toutes les prdp~rat ions enzymatiques,
l 'exception des ~-galactosidasiqucs et sur tout ~-glu- cosidasiques, peuvent se conserver h 4°C. Le pH opti- mum de chaque glycosidase a dtd ddtermin~ avec les o- ou p-ni t ropb6nyl-D-glyeopyranosides comme substrats .
BIBLIOGRAPHIE.
1. Conchie, J., Findlay, J. a Levvy, G. A. (1959) Bio- chem. J., 71, 318-325.
2. Findlay, J. ~ Levvy, G. A. (1960) Biochem. J., 77, 170-175.
3. Caygill, J. C., Rosto~, C. P. J. ~ Jevons, F. R. (1966) Biochem. J., 98, 405-409.
4. Watkins, W. M. (1959) Biochem. J., 71, 261-274. 5. Li, Y.-T., Li, S. C. ~ Shetlar, M. R. (1963) Arch.
Biochem. Biophys., 103, 436-442. 6. Hughes, R. C. ~ Jeanloz, R. W. (1964) Biochemistry,
3, 1543-1548. 7. Li, Y.-T. (1966) Y. Biol. Chem., 241, 1010-1012. 8. Petek, F. ~, Dong, T. (1961) Enzymot. , 23, 133-142. 9. Levvy, G. A. & McAllan, A. (1962) Nature, 195, 387.
10. Agrawal, K. M. L. ~ Bahl, O. P. (1968) J. Biol. Chem., 243, 103-111.
11. Neuberger, A. & Marshall, R. D. (1966) in Glyco- proteins (Gottschalk, A. ed.), BBA Library, vol. 5, p. 263, Elsevier Publ ishing Com~pany, Amster- dam.
12. Petek, F., Villarroya, E. ~ Courtois, J. E. (1969) European J. Biochem., 8, 395-402.
13. Gornall, A. G., BardawiI1, C. J. ~ David, M. M. (1949) J. Biol. Chem., 177, 751-766.
14. Lowry, O. H., Rosebrough, N. J., Farr, A. L. Randall , R. J. (1951) J. Biol. Chem., 193, 265-275.
15. Couchie, J. ~ Hay, A. J. (1959) Biochem. J., 73, 327- 334.
16. McIlvaine, T. C. (1921) J. Biol. Chem., 49, 183-186. 17. Boyd, W. C. (1963) Vox Sang., 8, 1-32. 18. Courtois, J. E. ~ Petek, F. (1966) Methods in Enzy-
mologg (Colo~vick, S. P. et Kaplau, N. O. ed.), vol. VIII, p. 565, Academic Press, New York and London.
19. Dey, P. M. & Pr idham, J. B. (1969) Biochem. J., 113, 49-55.
20. Dedonder, R. A. (1961) Annu. Reo. Biochem., 30, 347-382.
BIOCHIMIE, 1973, 55, n ° 1.





























