Embed Size (px)
Citation preview

Aus der Abteilung für Neurophysiologie
Leiter: Prof. Dr. Ulf Eysel
Ruhr-Universität Bochum
Neuroanatomische Untersuchungen zur
Immunhistochemie und den Verschaltungsmustern des
Nucleus subpraetectalis im visuellen System der Taube
(Columba livia)
Inaugural – Dissertation zur
Erlangung des Doktorgrades der Medizin
einer
Hohen Medizinischen Fakultät
der Ruhr-Universität Bochum
vorgelegt von
Marion Pilar Helene Theiß, geb. Meller aus Göttingen
2001

Dekan: Prof. Dr. med. Gert Muhr
Referent: Prof. Dr. med. Ulf Eysel
Korreferent: Prof. Dr. med. Walter Gehlen
Tag der mündlichen Prüfung: 07.02.2002

Inhaltsverzeichnis 2
1 EINLEITUNG 7
1.1 Fragestellung 8
1.2 Das visuelle System der Taube 9 1.2.1 Retina 10
1.2.1.1 Retinale Projektionen 11 1.2.2 Thalamofugales System 12 1.2.3 Tectofugales System 12
1.2.3.1 Tectum opticum 12 1.2.3.1 Nucleus rotundus 15
1.2.4 Praetectale Kerne 18 1.2.4.1 Nucleus subpraetectalis und Nucleus interstitio-praetecto-subpraetectalis 18 1.2.4.2 Konnektivitäten der praetectalen Kerne 18 1.2.4.3 Biochemische Charakterisierung der praetectalen Kerne 20 1.2.4.4 Mögliche Funktionen der praetectalen Kerne 22
1.3 Grundlagen der visuellen Systeme von Vögeln und Säugern 23
1.4 Neuronale Elemente der Signalübertragung 25 1.4.1 Ionotrophe Glutamat-Rezeptoren 26 1.4.2 Ionotrophe GABA-Rezeptoren 27 1.4.3 Glutamatdecarboxylase 28 1.4.4 Parvalbumin 28
1.5 Zielsetzung der Arbeit 29
2 MATERIAL UND METHODEN 30
2.1 Material 30
2.2 Tracing 30
2.3 Tracer 30 2.3.1 BDA 31 2.3.2 CtB 31
2.4 Tracingverfahren 31
2.5 Überlebenszeiten 32
2.6 Perfusion 32
2.7 Postfixieren 32
2.8 Schneiden der Gehirne 33
2.9 Darstellung von BDA 33
2.10 Darstellung von CtB 33
2.11 Auswertung 34
2.12 Immunhistochemie 34
2.13 Allgemeine Antikörperherstellung 34

Inhaltsverzeichnis
3
2.14 Eingesetzte Antikörper 35 2.14.1 Anti-Glutamatdecarboxylase 35 2.14.2 Anti-GABAA-Rezeptor (? -Untereinheit) 36 2.14.3 Anti-AMPA-Rezeptoren 36 2.14.4 Anti-Parvalbumin 36
2.15 Sekundäre Antikörper 36
2.16 Die Gewebeaufbereitung 37 2.16.1 Die Perfusion 37 2.16.2 Postfixieren 37 2.16.3 Gefrierschutz 38
2.17 Gewebebehandlungen - Immunreaktionen 38
2.18 ABC-Reaktion 39
2.19 DAB-Reaktion 39
2.20 Kontrollen 40
2.21 Färbung mit Kresylviolett 40
2.22 Eindeckeln 40
2.23 Auswertung 41 2.23.1 Qualitative Auswertungen 41 2.23.2 Quantitative Auswertungen 41
3 ERGEBNISSE 42
3.1 Tracing 42 3.1.1 BDA 45 3.1.2 CtB 56
3.2 Immunhistochemie 65 3.2.1 Kontrollexperimente 65 3.2.2 Qualitative Auswertung 65 3.2.3 Quantitative Auswertung 72
3.2.3.1 Quantitative Auswertung im Nucleus subpraetectalis 72 3.2.3.2 Quantitative Auswertung im Nucleus interstitio-praetecto-subpraetectalis 77
4 METHODENDISKUSSION 78
4.1 Injektionen von BDA und CtB 78
4.2 Immunhistochemie 79
5 VERSCHALTUNGSMUSTER DES NUCLEUS SUBPRAETECTALIS 81
5.1 Tectum opticum-Nucleus subpraetectalis 83
5.2 Nucleus subpraetectalis – Nucleus rotundus 84
5.3 Nucleus subpraetectalis-Nucleus triangularis 85
5.4 Nucleus subpraetectalis - Nucleus praetectalis 85

Inhaltsverzeichnis
4
6 BIOCHEMISCHE ORGANISATION DES SP/IPS-KOMPLEXES 86
7 INTEGRATION DES SP/IPS-KOMPLEXES IN DAS TECTOFUGALE SYSTEM 88
8 MÖGLICHE MECHANISMEN EINER PARALLELEN INFORMATIONSVERARBEITUNG 94
9 ZUSAMMENFASSUNG 96
10 LITERATURVERZEICHNIS 99

Abkürzungen
5
Abkürzungen AMPA ? -Amino-3-hydroxy-5-methyl-isoxazole-4-propriosäure
ÄnS Äußere nukleäre Schicht
ÄpS Äußere plexiforme Schicht
BDA Biotin-Dextran-Amin
CO Kohlenmonoxyd
CtB ? -Untereinheit des Choleratoxin
DAB-Reaktion Diaminobenzidin-Reaktion
DMSO Dimethylsulfoxid
GABA ? -Aminobuttersäure
GAD Glutamatdecarboxylase
Gld Nucleus geniculatus lateralis pars doralis
Glu R Glutamatrezeptor
GzS Ganglienzellschicht
H2O2 Wasserstoffperoxid
HRP horseradish peroxidase
IHC Immunhistochemie
Imc Nucleus isthmi, pars magnocellularis
InS Innere nukleäre Schicht
IpS Innere plexiforme Schicht
IPS Nucleus interstitio-praetecto-subpraetectalis
NMDA N-methyl-D-aspartat
NO Stickoxid
nPV Nucleus posteroventralis thalami (Kuhlenbeck)
nTT Nucleus tectothalamicus
Ov Nucleus ovoidalis
pAK Primärer Antikörper
PBS Phosphatpuffer
PT Nucleus praetectalis
PV Parvalbumin
RT Nucleus rotundus
Ru Nucleus ruber
SAC Stratum album centrale
sAK Sekundärer Antikörper

Abkürzungen
6
SGC Stratum griseum centrale
SGF Stratum griseum et fibrosum superficiale
SOp Stratum opticum
SP Nucleus subpraetectalis
SpL Nucleus spiriformis lateralis
T Nucleus triangularis
TO Tectum opticum
TrO Tractus opticus
TT Tractus tectothalamicus
V Ventrikel

Einleitung
7
1 Einleitung
Für Individuen, die aktiv mit ihrer Umgebung im Dialog stehen, sind Kenntnisse
über die Umgebung von höchster Bedeutung. Spezielle sensorische Systeme
nehmen bei den sich aktiv bewegenden Organismen Informationen aus der
Umwelt auf, welche anschließend mittels einer übergeordneten Steuereinheit
verarbeitet und in angemessener Weise beantwortet werden können. Die
Evolution der Organismen auf der Erde trieb die Entwicklung der zunächst
einfachen sensorischen Systeme immer weiter an, wobei auch die übergeordnete
Steuereinheit als Integrationsstelle dieser Informationen stetig weiterentwickelt
wurde. In der Gegenwart erreichten die phylogenetisch jungen Klassen der Vögel
und Säuger eine hohe Komplexität verschiedener sensorischer Systeme, zum
Beispiel des auditiven und visuellen Systems. Kennzeichnend für diese
verschiedenen Systeme, ist die selektive Perzeption und Verarbeitung einzelner
Stimuli der Umwelt, die über geeignete Rezeptoren aufgenommen und in
elektrische Signale umgewandelt werden, um anschließend vom Rezeptor in die
zentralen Bereiche der Steuereinheit geleitet zu werden. Als Steuereinheit fungiert
bei Vögeln und Säugern das zentrale Nervensystem, und für die integrative
Verarbeitung sensorischer Informationen als höchster Koordinationszentrum
insbesondere das Gehirn.
In den Neurowissenschaften Gegenstand intensiver Forschung ist das visuelle
System, welches mit seinen in der Retina des Auges gelegenen Rezeptoren
visuelle Informationen in elektrische Impulse umwandelt, die entlang
verschiedener Bahnen zu einzelnen Strukturen im Gehirn geleitet werden, um dort
ein Abbild der Umgebung zu erzeugen. Hierbei werden visuelle Stimuli gemäß
ihrer visuellen Information in unterschiedlichen Bahnen des Sehsystems
verarbeitet, wobei die Informationen bereits auf retinaler Ebene gefiltert und
aufgetrennt werden. Diese Filterung der Information ist ein Prozeß, bei dem
morphologische, elektrophysiologische und biochemische Eigenschaften der
beteiligten neuronalen Systeme den Informationsgehalt regulieren. So finden sich
in der Retina der Vertebraten Neuronen, die vornehmlich auf bewegte Stimuli
reagieren, während andere Neuronen Detailinformation der visuellen Szenerie
perzipieren. Desweiteren sind zum Beispiel bestimmte Photorezeptoren der
Retina, die Stäbchenzellen, selektiv erregbar durch Helligkeitsunterschiede,

Einleitung
8
wiederum andere Photorezeptoren, die Zapfenzellen, nehmen hingegen
Farbinformationen der Umwelt war.
Die Verarbeitung visueller Informationen in parallelen aufsteigenden Bahnen von
der Retina zu zentralen Kerngebieten des Gehirns legt die Vermutung nahe, daß
jede dieser Projektionen aus der Retina auch spezielle Funktionen bei der
Verarbeitung visueller Informationen erfüllt. Hierbei bestimmen vermutlich bereits
die Projektionen verschiedener retinaler Neuronen, die zum Beispiel
Bewegungsinformationen oder Detailinformationen der visuellen Umwelt
weiterleiten, den visuellen Informationsgehalt in den entsprechenden visuellen
Sehbahnen. Gleichwohl sind auch zwischen den aufsteigenden Sehbahnen
Verschaltungen bekannt, die eine wechselseitige Modulation zunächst distinkter
Informationen wahrscheinlich werden lassen.
1.1 Fragestellung
Das visuelle System der Vertebraten gehört im Nervensystem zu den bislang am
besten charakterisierten sensorischen System, wobei verschiedene zum visuellen
System gehörende Hirnstrukturen in ihrer Funktion erfaßt sind. Dennoch sind viele
Wege der Informationsfortleitung und Informationsverarbeitung innerhalb dieses
Sehsystems bei den Vertebraten auch gegenwärtig noch nicht komplex erfaßt.
Weitere Erkenntnisse über die Verarbeitung visueller Informationen können
morphologische und biochemische Untersuchungen der aufsteigenden
Sehbahnen, beziehungsweise der beteiligten neuronalen Strukturen liefern. Da die
Funktionen einzelner Abschnitte in einem sensorischen System durch die
morphologischen und biochemischen Möglichkeiten / Eigenschaften bestimmt
sind, lassen zum Beispiel Untersuchungen der Morphologie von Neuronen, der
Verknüpfungen einzelner Hirnstrukturen sowie der Verteilung von
Neurotransmittern in einzelnen Abschnitten des Gehirn Rückschlüsse auf die
Informationsverarbeitung in dem jeweiligen Bereich zu.
In der vorliegenden Arbeit werden anatomische und biochemische Eigenschaften
praetectaler Kerne untersucht, die möglicherweise an der visuellen
Informationsverarbeitung, bei der Taube Columba livia, beteiligt sind. Das visuelle
System der Taube kennzeichnet zwei aufsteigenden Bahnen, die tectofugale und
die thalamofugale Sehbahn, die mit dem genikulostriatalen beziehungsweise
extragenikulären System bei Säugern verglichen werden (Karten & Shimizu,

Einleitung
9
1989). Das in der vorliegenden Arbeit untersuchte Nucleus
subpraetectalis/Nucleus interstitio-praetecto-subpraetectalis-Komplex (SP/IPS-
Komplex) steht in enger anatomischer Lokalisation zum tectofugalen System. Eine
mögliche Funktion dieses Komplexes ist eine Modulation der tectofugalen
Informationsverarbeitung. Hierbei stellt sich die Frage in welchem Umfang das
tectofugale System modulatorische Einflüsse von anderen Systemen erhält. Um
die Hypothese zu unterstützen, daß der SP/IPS-Komplex einen anatomischen
Bezug zum tectofugalen System hat, wurden die Konnektivitäten des SP/IPS-
Komplexes durch Injektion von retrograden und anterograden Tracer in den
SP/IPS untersucht.
Im zweiten Teil der Arbeit wurden mögliche biochemische Signalwege des
SP/IPS-Komplexes untersucht, daß heißt mögliche intranukleäre
Informationsvermittlungen, sowie efferente Projektionen anhand des
Expressionsmusters inhibitorischer und exzitatorischer Neurotransmitter und
Rezeptortypen erfaßt.
Im Folgenden wird zunächst das visuelle System der Taube Columba livia
vorgestellt, mit einem Schwerpunkt auf dem asymmetrisch organisierten
tectofugalen System, bevor mögliche Funktionen der praetectalen Kerne skizziert
werden.
1.2 Das visuelle System der Taube
Die Taube ist eine visuell hochentwickelte Spezies (Emmerton, 1983). Visuelle
Informationsverarbeitung beginnt mit der Aufnahmen des Lichtreizes im Auge. Das
in der Linse und im Glaskörper gebrochene Licht wird in den Photorezeptoren der
Retina in ein elektrisches Spannungssignal umgewandelt, wodurch die visuelle
Information über diverse retinale Verschaltungen schließlich im Nervus opticus zu
weiteren neuronalen Strukturen geleitet wird. Bei der Taube sind insbesondere
Hirnstrukturen der beiden großen aufsteigenden Sehbahnen an visueller
Informationsverarbeitung beteiligt. Zum einen ist es das thalamofugale System,
dessen Ursprung der Nucleus geniculatus lateralis, pars dorsalis ist, welcher zum
visuellen Wulst im Vorderhirn projiziert, und zum anderen das tectofugale System,
mit den Projektionen vom Tectum opticum zum diencephalen Nucleus rotundus
und schließlich zum telencephalen Ectostriatum.

Einleitung
10
1.2.1 Retina
Die sechsschichtige Retina der Taube zeigt einen für Vertebraten typischen
Aufbau aus zellkörperreichen (nukleären) und zellkörperarmen (plexiformen)
Schichten. Bei der adulten Taube werden zwei plexiforme Schichten, eine innere
plexiforme (IpS) und eine äußere plexiforme (ÄpS) sowie drei nukleäre Schichten,
die sich in eine innere nukleäre (InS) und äußere nukleäre (ÄnS) und eine
Ganglienzellschicht (GzS) gliedern, unterschieden. Die Photorezeptoren befinden
sich in der ÄnS, während die Horizontalzellen, Amakrinzellen, die inter-plexiformen
Zellen und die Bipolarzellen in der InS lokalisiert sind. Die Ganglienzellen sind
perlschnurartig nebeneinander in der entsprechenden GzS angeordnet (Wässle &
Boycott, 1991).
In der ÄnS werden zwei Typen von Photorezeptoren unterschieden: erstens, die
für das Dämmerungssehen verantwortlichen Stäbchen, und zweitens, die für das
Farbensehen wichtigen Zapfen und Doppelzapfen (Morris, 1970; Mariani & Leure-
DuPree, 1978). Gemeinsam ist beiden Photorezeptortypen der Aufbau aus einem
inneren und einem äußeren Segment wobei das innere Segment in der ÄnS und
das äußere Segment in der ÄpS lokalisiert ist (Nalbach et al., 1993). Das innere
Segment dient der Energieversorgung, während das äußere Segment den
eintreffenden Lichtreiz in ein elektrisches Spannungssignal umwandelt. Das
elektrische Signal gelangt anschließend weiter über die Bipolarzellen zu den
Ganglienzellen, deren Axone als einzige die Retina verlassen und den Nervus
opticus bilden.
Die Retina der Taube wird unterteilt in ein rotes Feld im dorso-temporalen Bereich,
und ein gelbes Feld, welches die übrige Retina bestimmt. Die Namensgebung
dieser Felder beruht auf farbigen Öltropfen im inneren Segment der Zapfen (Bloch
& Maturana, 1971; Mariani & Leure-DuPree, 1978). Diese Öltropfen setzen sich
aus Lipiden und Karotinoiden zusammen, deren Konzentration die Farbe der
Photorezeptoren bestimmt (Varela et al., 1993). Galifret (1968) stellte fest, daß im
roten Feld der Retina rote und orangene Öltropfen überwiegen sowie im gelben
Feld gelbe und grüne Öltropfen. Die Aufgaben der Öltropfen sind einerseits eine
Filterfunktion zum Schutz der Retina vor UV-Strahlung (Emmerton, 1983), und
andererseits die Absorption des Lichtes eines speziellen Wellenlängenbereiches,

Einleitung
11
um eine maximale Aktivierung des jeweiligen Photorezeptors zu erreichen (Young
& Martin, 1984).
Im roten und gelben Feld der Retina können desweiteren Bereiche mit erhöhter
Ganglienzelldichte abgegrenzt werden, die als Area dorsalis im roten Feld, und als
Area centralis im gelben Feld bezeichnet werden (Bingelli & Paule, 1969; Hayes &
Holden, 1983). Mit diesen morphologischen Unterscheidungskriterien geht parallel
auch eine unterschiedliche visuelle Informationsverarbeitung einher. Im dorso-
temporal gelegenen roten Feld werden vornehmlich frontale, binokulare Reize
abgebildet, die das Erkennen von nahen Objekten ermöglichen (Bloch &
Martinoya, 1982; Uhlrich et al. 1982). Das gelbe Feld ist dagegen verantwortlich
für das laterale, monokulare Gesichtsfeld. Blough (1971) stellte in
Verhaltensexperimenten fest, daß die Fixierung weit entfernter Objekte mit der
Area centralis des gelben Feldes erfolgt. Hahmann und Güntürkün (1993) zeigten
darüber hinaus in anatomischen Untersuchungen und Verhaltensexperimenten,
daß eine maximale visuelle Auflösung in der Area centralis vorliegt.
Eine Besonderheit der Taubenretina ist die schon auf retinaler Ebene
stattfindende komplexe Reizverarbeitung, wobei in elektrophysiologischen
Untersuchungen die Neuronen auf Bewegung, Orientierung oder Helligkeit selektiv
reagierten (Maturana 1962; Maturana & Frenk 1963). Diese Form der
Reizverarbeitung findet man bei Säugern erst in nachgeschalteten Hirnarealen.
1.2.1.1 Retinale Projektionen
Der Nervus opticus der Taube besteht aus ca. 2,4 Millionen Axone, das heißt dem
2,5 fachen des Nervus opticus beim Menschen (Curcio & Allen 1990). Bei den
Tauben kreuzt der Sehnerv nahezu vollständig im Chiasma opticum (Cowan et al.
1961), mit nur geringen ipsilateralen Projektionen zum Nucleus of the Basal Optic
Root (nBor) (Güntürkün & Karten, 1991). Shimizu (1994) bestätigt diese Daten,
und demonstrierte weitere ipsilaterale Verbindungen zum thalamischen Nucleus
geniculatus lateralis pars ventralis, zu den beiden praetectalen Kernen Nucleus
praetectalis parvocellularis und magnocellularis, wie auch zum Nucleus lateralis
anterior. Letztere Projektionen wurden durch eine Arbeit von Remy und Güntürkün
(1991) bestätigt.
Der Großteil der retinalen Projektionen kreuzt im Chiasma opticum zur
kontralateralen Hirnhälfte, und erreicht dort den Nucleus geniculatus lateralis pars

Einleitung
12
ventralis, den Nucleus suprachiasmaticus, die Area praetectalis, den Nucleus
lentiformis mesencephali, den Nucleus of the Basal Optic Root, den Nucleus
geniculatus lateralis pars doralis (Gld) und das Tectum opticum. Der Nucleus
geniculatus lateralis pars dorsalis und das Tectum opticum bilden den
Ausgangspunkt für das thalamofugale und das tectofugale System, die beiden
großen aufsteigenden Sehbahnen der Taube.
1.2.2 Thalamofugales System
Das thalamofugale System stellt die Verbindung zwischen den Nucleus
geniculatus lateralis, pars dorsalis (Gld) und dem visuellen Wulst im
Telencephalon dar. Der visuelle Wulst erhält bilaterale Eingänge des Gld. Eine
vergleichbare visuelle Sehbahn bei Säugern stellt das genikulostriatäre System
dar, wobei hier der Nucleus geniculatus lateralis pars dorsalis auf den visuellen
Cortex projiziert (Karten & Shimizu, 1989).
1.2.3 Tectofugales System
Das tectofugale System bildet neben dem thalamofugalen System die weitere zum
Telencephalon aufsteigende Sehbahn, deren Ausgangspunkt das Tectum opticum
bildet. Das Tectum opticum projiziert bilateral zum diencephalen Nucleus rotundus
(Hellmann & Güntürkün, 1999), und dessen Efferenzen erreichen das ipsilaterale
telencephale Ectostriatum. Das tectofugale System der Vögel wird mit dem
extragenikulären System der Säuger verglichen, bei denen die Projektionen über
den mesencephalen Colliculus superior zum thalamischen Pulvinar, und von dort
letztendlich zum extrastriatalen Cortex gelangen (Karten & Shimizu, 1989).
1.1.1.1 Tectum opticum
Das Tectum opticum der Taube liegt im Mesencephalon. Zirka 75-95% der
retinalen Efferenzen kreuzen im Chiasma opticum und gelangen zum
kontralateralen Tectum opticum (Remy & Güntürkün, 1991). Die Morphologie des
Tectum opticum zeigt sich primär durch eine laminäre Strukturierung, die
abwechselnd aus nukleären und plexiformen Schichten besteht. Ramón y Cajal
(1911) unterteilte das Tectum opticum in fünfzehn Schichten, wobei die Schicht 1

Einleitung
13
pial lokalisiert ist und die Schicht 15 an den Seitenventrikel angrenzt. Weitere
morphologische Charakterisierung wurden durch die Definition verschiedener
Zelltypen (Hayes & Webster, 1975), Unterschiede in der dorso-ventralen
Schichtdicke des Tectum opticum (Hayes & Webster, 1985), und Kontraste in der
Zelldichte (Theiss et al., 1998) festgestellt.
Die retinale Innervation des Tectum opticum ist topographisch organisiert, und
terminiert in den superficialen Schichten 2 bis 7 (McGill et al., 1966; Crossland et
al., 1974; Clark & Whitteridge, 1976; Remy & Güntürkün, 1991). In diesen
retinorezipienten Schichten enthalten vor allem die Schichten 3, 5 und 7 retinale
Terminalien (Hayes & Webster, 1975; Angaut & Repérant, 1978; Acheson et al.,
1980).
Die Zelltypen in den superficialen Schichten sind die Horizontalzellen und
Radialzellen. Während die Horizontalzellen durch in den Schichten 4 und 5
gelegene Perikaria und parallel zur tectalen Oberfläche verlaufenden Dendriten
charakterisiert sind, liegen die Somata der Radialzellen auch in anderen
superficialen Schichten, und ihre Dendriten ziehen durch mehrere tectale
Schichten radial zur Oberfläche. Funktionell erhalten die Horizontalzellen direkten
retinalen Eingang, projizieren aber nicht aus dem Tectum opticum heraus (Van
Gehuchten, 1892), sondern fungieren als Interneuronen innerhalb dieser
mesencephalen Struktur. Die Funktion der in den superficialen tectalen Schichten
gelegenen Radialzellen ist vermutlich ähnlich, wobei aber Projektionen aus dem
Tectum opticum heraus nicht ausgeschlossen werden können.
In den mittleren Zellschichten des Tectum opticum, in den Schichten 8 bis 12, sind
Radialzellen und multipolare Neuronen lokalisiert (Van Gehuchten, 1892). Die
Dendriten dieser Nervenzellen steigen bis in die superficialen Schichten auf und
haben dort synaptischen Kontakt mit den retinalen Ganglienzell-Axonen oder den
Horizontalzellen (Van Gehuchten, 1892; Ramon y Cajal, 1911; Hardy et al., 1985).
Efferenzen der Radialzellen ziehen zum ventralen Thalamus und dem Nucleus
geniculatus lateralis pars dorsalis (Hunt & Künzle, 1976; Wild, 1989). Die
Efferenzen der multipolaren Neuronen gelangen vermutlich zu den praetectalen
Kernen.
In der tiefen tectalen Schicht 13 sind die großen multipolaren Neuronen lokalisiert,
welche mit ihren Efferenzen den Hauptanteil des Informationsausganges des
Tectum opticums bilden, und vor allem zum ipsi- wie kontralateralen Nucleus

Einleitung
14
rotundus projizieren. Diese tecto-rotundalen Projektionen zeigen auffällige dorso-
ventral Unterschiede in der Faserdichte, abhängig vom Ursprung im Tectum
opticum, wobei das ventrale Tectum opticum mit mehr Efferenzen zum Nucleus
rotundus projiziert als das dorsale Tectum opticum (Hellmann & Güntürkün, 1999).
Periventrikulär lokalisiert sind die Schichten vierzehn und fünfzehn, bei denen es
sich um reine Faserschichten handelt.
Elektrophysiologische, autoradiographische und immunhistochemische
Untersuchungen führten zu weiteren Kenntnissen über die Funktionalitäten der
verschiedenen Zelltypen im Tectum opticum. Karten und Mitarbeiter (1997)
unterscheiden anhand der Lokalisation und Ausbreitung zwei multipolare
Neuronentypen in der Schicht 13. Typ I der SGC-Neurone sind in der
Außenschicht lokalisiert und erhalten ihren tectalen Input aus der Schicht 5b.
Typisch für die Typ I Neurone ist ihre ausgeprägte bürstenartige Aufzweigung. Die
Typ II Neurone des SGC befinden sich dagegen in der Innenschicht. Der
Dendritenbaum der Typ II Neurone reicht jedoch nur bis Schicht 8 und zeigt einen
elliptischen Aufbau. Bei Hühnern wird von Luksch und Mitarbeitern (1998) ein
weiterer Neuronentyp im SGC beschrieben. Kennzeichen für die Typ III Neuronen
des SGC sind lange dendritische Felder, die in Schicht 4 des Tectum opticum
terminieren. Hellmann und Güntürkün (2001) beschreiben nach Tracerinjektionen
in den Nucleus rotundus fünf unterschiedliche Neuronentypen, Typ I bis V, in der
Schicht 13 des Tectum opticum. Folgende Unterscheidungskriterien wurden
gewählt: die sublaminäre Lokalisation in der Schicht 13, die Somatagröße,
Projektionen zu unterschiedlichen Subarealen des Nucleus rotundus und des
Nucleus triangularis und Differenzen innerhalb der Verbindungen des Tectum
opticum zu den retinorezipienten Schichten 3 bis 7.
Hardy und Mitarbeiter (1985) untersuchten die intratectale visuelle
Informationsverarbeitung, wobei sie zeigten, daß die multipolaren Neuronen der
Schicht 13 direkten retinalen Input erhalten, und diesen ohne synaptische
Verschaltungen weiterleiten. Auch die Radialzellen der Schichten 10 und 11 sind
direkt mit retinalen Zellen verschaltet, wobei diese aber vermutlich als
Interneuronen agieren um die elektrische Erregung zu den Neuronen der Schicht
13 weiterzuleiten, wo sie axo-somatische Synapsen bilden. Autoradiographische
Experimente deuten darauf hin, daß Glutamat als exzitatorischer Transmitter für
die Signalfortleitung im Tectum opticum verantwortlich ist (Beart, 1976; Henke et

Einleitung
15
al.,1976a/b). Immunhistochemische Untersuchungen zur Expression des
Glutamatrezeptors vom AMPA-Typ im Tectum opticum bestätigten diese
Annahme, und zeigten die Expression von Glutamatrezeptoren in den
superficialen Schichten, in den Schichten 9 und 10 und intensiv in der Schicht 13,
wodurch glutamaterge Mechanismen bei der retino-tectalen Signalfortleitung als
auch bei der intratectalen Signalfortleitung wahrscheinlich sind (Theiss et al.,
1998).
1.2.3.1 Nucleus rotundus Der Nucleus rotundus ist der größte diencephale Kern im Taubenhirn und bildet
nach dem Tectum opticum die folgende Verarbeitungsebene der visuellen
Informationen im tectofugalen System. Bei der morphologischen Charakterisierung
des Nucleus rotundus zeigt sich ein homogener Aufbau. Tömböl und Mitarbeiter
(1992) unterschieden in Golgi-Studien mit Hühnern zwei Neuronentypen aufgrund
ihrer Größe und der Verzweigung der Dendritenbäume. Die großen Neuronen
imponieren durch weitreichende Dendriten und zum Ectostriatum projizierende
Axone, während die kleinen Neuronen vermutlich als Interneuronen mit
modulatorischen Aufgaben fungieren. Eine zumeist inhibitorische
Signalmodulation der kleineren Neuronen wird durch ihre Terminalien mit zumeist
symmetrischen Synapsen auf den Perikarien und Dendriten der großen Zellen
angenommen (Tömböl et al., 1992). Exzitatorische Signale resultieren nach
Tömböl vermutlich aus den einziehenden tectalen Fasern, die asymmetrische
Synapsen mit Glomeruli-ähnliche Strukturen erkennen lassen.
Der Nucleus rotundus erhält Eingänge aus dem Tectum opticum, wobei die
multipolaren Zellen der Schicht 13 des Tectum opticum bilateral zum Nucleus
rotundus und zum Nucleus triangularis, der dem Nucleus rotundus dorsal
aufliegenden Kappe, projizieren (Karten & Revzin, 1966; Karten & Hodos, 1970;
Benowitz & Karten, 1976; Bischof & Niemann, 1990; Güntürkün et al., 1993;
Hellmann & Güntürkün, 1999). Diese Afferenzen ziehen vor allem in den ventralen
Nucleus rotundus ein (Granda & Yazulla, 1971). Güntürkün und Melsbach (1992)
stellten Unterschiede in den kontralateralen Projektionen beider Hemisphären fest,
wobei der linke Nucleus rotundus massiven Eingang vom rechten Tectum opticum
erhält, während die Projektionen vom linken Tectum opticum zum rechten Nucleus
rotundus in der Anzahl deutlich schwächer sind. Benowitz und Karten (1976)

Einleitung
16
beschreiben eine topographische Organisation der Efferenzen des Tectum
opticum zum Nucleus rotundus, bei der die oberflächlichen multipolaren Zellen der
Schicht 13 in den rostralen Bereich des Nucleus rotundus projizieren, und der
mediale und caudale Nucleus rotundus Eingänge aus tiefer gelegenen Zellen der
Schicht 13 erhält. Diese topographische Organisation wurde durch Nixdorf und
Bischof (1982) bei Zebrafinken bestätigt. Weitere Tracinguntersuchungen zur
topographischen Organisation der tectorotundalen Projektion wurden durch Karten
und Mitarbeiter (1997) und Deng und Rogers (1998) durchgeführt. Karten injizierte
einen Tracer in verschiedene Bereiche des Nucleus rotundus und konnte
retrograd zwei Neuronentypen mit unterschiedlicher Projektion in der Schicht 13
des Tectum opticum feststellen. Markierung der Außenschicht der Schicht 13
fanden sich bei Tracerapplikation in den rostralen und zentralen Nucleus rotundus,
während Injektionen in den caudalen Nucleus rotundus und in den Nucleus
triangularis Neuronen der inneren Schicht 13 (zur Schicht 14 gewandt) darstellten.
Deng und Rogers (1998) beschrieben ebenso unterschiedliche Projektionsziele
innerhalb der Schicht 13. Der innerste Anteil, übergehend zur Schicht 14, projiziert
demnach zum Nucleus triangularis. Die mittlere Schicht 13 zeigt hingegen
Projektionen zum dorsalen Nucleus rotundus, und Neuronen im äußeren Anteil
der Schicht 13 projizieren zum ventralen Nucleus rotundus.
Efferenzen des Nucleus rotundus gelangen zum ipsilateralen Ectostriatum,
welches im Telencephalon lokalisiert ist (Kondo, 1933; Karten & Hodos, 1970;
Benowitz & Karten, 1976; Nixdorf & Bischof, 1982; Miceli & Repérant, 1985). Bei
Hühnern entspringen diese Projektionen ausschließlich von den großen Neuronen
des Nucleus rotundus (Tömböl, 1992).
Um den Nucleus rotundus biochemisch zu charakterisieren wurden diverse
Untersuchungen über die Verteilung verschiedener Proteine, Neurotransmitter und
Rezeptoren durchgeführt. Braun und Mitarbeiter (1985) untersuchten die
Verteilung des Calcium-bindenden Proteins Parvalbumin sowie des
Aktivitätsmarkers 14C-Desoxyglucose unter Lichteinfluß bei Zebrafinken, und
fanden eine homogene Verteilung im gesamten Kern. Ebenso wurde eine
schwache, aber gleichmäßige Verteilung von GABA-positiven Neuronen
beobachtet (Domenici et al., 1988, Ngo et al., 1992). Hingegen konnten weder
muscarinerge noch nikotinerge Acetylcholinrezeptoren im Nucleus rotundus

Einleitung
17
detektiert werden (Wachtler, 1985; Dietl et al., 1988a), obwohl Martinez de la
Torre (1990) bei Hühnern der Nachweis von Acetylcholinesterase in dieser
Hirnstruktur gelang.
Bei der Bestimmung der Größe der rezeptiven Felder rotundaler Neuronen stellte
man fest, daß 80% der Neuronen im Nucleus rotundus mit 100° bis 175° ähnlich
große rezeptive Felder besitzen, wie die Neuronen in den tiefen tectalen Schichten
(Revzin, 1979; Maxwell & Granda, 1979). Mit Hilfe elektrophysiologischer Arbeiten
wurde der Nucleus rotundus und der Nucleus triangularis in verschiedenen
Untersuchungen in diverse funktionelle Bereiche unterteilt. Revzin (1979)
unterscheidet die drei Bereiche posterior, anterior und ventral; Zellen im
posterioren Drittel reagieren demnach vornehmlich auf bewegte Stimuli ohne
Vorzugsrichtung, während Zellen im anterioren Drittel bevorzugt durch
Reizmerkmale wie Größe, Richtung und Geschwindigkeit eines Objektes stimuliert
werden können. Neuronen der ventralen Region antworten hingegen vor allem auf
Helligkeitsunterschiede im Gesichtsfeld. Granda und Yazulla (1971) postulierten
nach elektrophysiologischen Ableitungen im Diencephalon der Taube, daß im
dorsalen Nucleus rotundus eher exzitatorische Signalmodulation und im ventralen
Nucleus rotundus zumeist inhibitorische Signalmodulation nach visueller
Stimulation erfolgt. Wang und Frost (1992) vermuten auf Grund
elektrophysiologischer Ableitungen, daß Neuronen im Nucleus rotundus „time to
contact“ Berechnungen durchführen. Diese sind wichtig, um Kollisionen mit
anderen Flugobjekten oder ruhenden Hindernissen voraus zu berechnen,
beziehungsweise zu vermeiden. Eine umfassende Arbeit von Wang und
Mitarbeiter (1993) teilt den Nucleus rotundus in vier funktionelle Bereich ein. Im
anterioren Nucleus rotundus werden dabei von dorsal nach ventral Bereiche
unterschieden, deren Neuronen auf Farbe, Helligkeit und Bewegung im
zweidimensionalen Raum reagieren. Neuronen des posterioren Nucleus rotundus
antworten im dorsalen Abschnitt vornehmlich auf Bewegung im dreidimensionalen
Raum, und im ventralen Bereich bevorzugt auf Bewegung im zweidimensionalen
Raum. Elektrophysiologische Untersuchungen von Huang und Mitarbeitern (1998)
zeigen, daß exzitatorische Signale aus dem ipsi- und kontralateralen Tectum
opticum im Nucleus rotundus durch den Neurotransmitter GABA eliminiert werden
können. Hierbei zeigt sich desweiteren eine generelle Hemmung von Neuronen
des Nucleus rotundus auf visuelle Reize. Auf Grund dieser Daten ist eine Funktion

Einleitung
18
von GABA als inhibitorischer Neurotransmitter im Nucleus rotundus anzunehmen,
wenngleich bislang ein direkter Nachweis von GABA und GABA-Rezeptoren nicht
gegeben ist.
1.2.4 Praetectale Kerne
Die Praetectalen Kerne umfassen den Nucleus praetectalis, Nucleus
subpraetectalis und den Nucleus interstitio-praetecto-subpraetectalis. Der Nucleus
subpraetectalis und der Nucleus interstitio-praetecto-subpraetectalis werden von
vielen Autoren als SP/IPS-Komplex zusammengefaßt. Da die Funktion des
Nucleus interstitio-praetecto-subpraetectalis noch weitgehend unbekannt ist, wird
dieser Kern gemeinsam mit dem Nucleus subpraetectalis aufgeführt.
1.2.4.1 Nucleus subpraetectalis und
Nucleus interstitio-praetecto-subpraetectalis
In der vorliegenden Arbeit werden die Einflüsse und die Funktionen des Nucleus
subpraetectalis und des Nucleus interstitio-praetecto-subpraetectalis untersucht.
Der zu den praetectalen Kernen zählende Nucleus subpraetectalis ist medial vom
Tectum opticum und ventro-lateral vom Nucleus rotundus im Taubenhirn
lokalisiert. Die Ausdehnung des Nucleus subpraetectalis in der caudo-rostralen
Ebene beginnt bei A4.25mm und reicht bis A5.25mm (Karten & Hodos, A
Stereotaxic Atlas of the Brain of the pigeon, 1967). Der Nucleus interstitio-
praetecto-subpraetectalis liegt dem Nucleus subpraetectalis als dorsale Kappe
auf, wobei seine caudo-rostrale Ausdehnung von A4.50mm bis A5.00mm reicht.
Die Zuordnung der beiden Kerne zu einer spezifischen Hirnregion variiert in der
Literatur. Benowitz und Karten (1976) rechnen den Nucleus subpraetectalis den
posterioren ventro-lateralen Diencephalon zu, während Domenici und Mitarbeiter
(1988) den Nucleus subpraetectalis dem Mesencephalon zuordnen.
1.2.4.2 Konnektivitäten der praetectalen Kerne
Für den Nucleus subpraetectalis wird eine inhibitorische Wirkung auf den Nucleus
rotundus vermutet (Benowitz & Karten, 1976; Shimizu et al., 1988; Mpodozis et al.,

Einleitung
19
1996; Deng & Rogers, 1998;). Die Rolle des Nucleus interstitio-praetecto-
subpraetectalis bleibt jedoch bislang weitgehend unklar.
Afferent verbunden sind der Nucleus subpraetectalis und der Nucleus interstitio-
praetecto-subpraetectalis der Taube mit dem Tectum opticum beider Hemisphären
(Hunt & Künzle, 1976a). Bei Zebrafinken wurden die Projektionen des Tectum
opticum mittels radioaktiv markierter anterograder Tracer untersucht. Der Nucleus
subpraetectalis und der Nucleus interstitio-praetecto-subpraetectalis, zusammen
als SP/IPS-Komplex beschrieben, erhalten demnach bilateralen tectalen Eingang
(Bischof & Niemann, 1990).
Um die Afferenzen des Nucleus rotundus näher zu untersuchen, injizierten
Benowitz und Karten (1976) Meerrettichperoxidase (horseradish peroxidase; HRP)
in definierte Abschnitte des Nucleus rotundus der Taube. Nach der Injektion in den
ventralen Nucleus rotundus stellten sie Markierungen ausschließlich in den
praetectalen Kernen Nucleus subpraetectalis und Nucleus interstitio-praetecto-
subpraetectalis fest. Wurde dagegen in den anteromedialen Nucleus rotundus
HRP appliziert, so ergaben sich zusätzlich zu den beiden praetectalen Kernen
auch somatische Markierungen im Tectum opticum. Nach HRP-Injektionen in den
caudalen Nucleus rotundus wurden jedoch keine Markierung im Nucleus
subpraetectalis oder Nucleus interstitio-praetecto-subpraetectalis beobachtet.
Benowitz und Karten resümierten anhand dieser Daten, daß der ventrale Nucleus
rotundus seine Afferenzen über den Nucleus subpraetectalis und Nucleus
interstitio-praetecto-subpraetectalis erhält. Außerdem vermuten sie den Nucleus
subpraetectalis als eine Zellansammlung hemmender Neuronen auf weitere
Kerngebiete.
Weitere Efferenzen des SP/IPS-Komplexes wurden durch HRP-Injektionen in den
telencephalen visuellen Wulst und das Ectostriatum bei Zebrafinken beschrieben
(Nixdorf & Bischof, 1982). Bei Applikation in das caudale Ectostriatum erkannten
die Autoren bei einer Injektion Markierungen im Nucleus subpraetectalis und
Nucleus interstitio-praetecto-subpraetectalis.
Tömböl und Mitarbeiter (1994) suchten den Ursprung einer inhibierenden
Innervation des thalamischen Nucleus rotundus bei Hühnern. Sie injizierten einen
Tracer in den Nucleus subpraetectalis und den Nucleus posterior ventralis thalami,
und konnten anschließend immunologisch GABAerge Strukturen an den
markierten Projektionen dieser beiden praetectalen Kerne im Nucleus rotundus

Einleitung
20
nachweisen. Im Nucleus rotundus wurden Fasern und Terminalien im gesamten
Kerngebiet beobachtet, die in ihrer Faserdichte variierten und keine
topographische Organisation erkennen ließen. In den immunologischen
Untersuchungen waren die durch den Tracer markierten Fasern zusätzlich GABA-
positiv. Deng und Rogers (1998) stellten eine topographische Organisation der
Afferenzen des Nucleus rotundus bei Hühnern nach Injektion eines retrograden
Tracers in distinkte Abschnitte des Nucleus rotundus fest. Im Gegensatz zu
Benowitz und Karten zeigten Deng und Rogers, das beim Huhn der gesamte
Nucleus rotundus Afferenzen aus dem Nucleus subpraetectalis, dem Nucleus
interstitio-praetecto-subpraetectalis und dem glutamatergen Tectum opticum
erhält. Der Nucleus subpraetectalis und der Nucleus interstitio-praetecto-
subpraetectalis projizieren demnach nur zum ipsilateralen Nucleus rotundus,
wobei der dorsale Nucleus subpraetectalis zum dorsalen Nucleus rotundus, der
ventrale Nucleus subpraetectalis zum ventralen Nucleus rotundus, und der
zentrale Nucleus subpraetectalis zum zentralen Nucleus rotundus projiziert. Nach
Deng und Rogers (1998) besteht eine Verbindung zwischen dem Nucleus
interstitio-praetecto-subpraetectalis und dem Nucleus triangularis. Außerdem
zeigten diese Autoren, daß im Nucleus subpraetectalis vornehmlich GABAerge
Neuronen lokalisiert sind.
1.2.4.3 Biochemische Charakterisierung der praetectalen Kerne
Um die Funktion und den Einfluß der praetectalen Kerne weiter zu verstehen,
müssen die biochemischen Charakteristika dieser Kerne näher untersucht werden.
Domenici und Mitarbeiter (1988) prüften die immunhistochemische Verteilung
GABAerger Neuronen im Taubenhirn. Im Nucleus subpraetectalis stellten sie eine
hohe Dichte GABA-positiver Somata fest, während gleichzeitig die Dichte eines
GABAergen Neuropils im Nucleus subpraetectalis deutlich geringer war. Hierbei
imponierten zwei Zellgrößen im Nucleus subpraetectalis: der mittlere Zelltyp mit
einer Somagröße von 15-20µm, und der große Zelltyp mit mehr als 20µm
Somagröße. Domenici und Mitarbeiter sehen den Nucleus subpraetectalis
aufgrund seiner GABAergen Neuronen als geeigneten Kandidat für den Ursprung
eines GABAergen Projektionssystem. Die Verteilung GABAerger Strukturen im
Nucleus rotundus und Nucleus subpraetectalis wurde von Shimizu und Mitarbeiter
(1988) vor und nach elektrolytischer Läsion des Nucleus subpraetectalis

Einleitung
21
beschrieben. Sie stellten fest, daß im Nucleus rotundus nach der Läsion des
Nucleus rotundus GABAerge Markierung verschwanden.
Granda und Crossland (1989) untersuchten immunologisch die GABAerge
Verteilung im Diencephalon und Mesencephalon von Hühnern kurz nach dem
Schlupf. Anschließend verglichen sie ihre Ergebnisse mit der GABAergen
Verteilung bei Säugern. Der Nucleus subpraetectalis und der Nucleus interstitio-
praetecto-subpraetectalis imponieren durch markierte Neurone, mit einer
Zellgröße von mehr als 100µm2. Die Zahl der GABA-positiven Fasern ist im
Nucleus subpraetectalis gering, wogegen im Nucleus interstitio-praetecto-
subpraetectalis sich zahlreiche GABA-positive Fasern darstellen. Die Autoren
vermuten, daß die großen GABA-positiven Neurone einen modulierenden Einfluß
auf exzitatorische Eingänge haben könnten.
Ngo beschrieb mit seinen Mitarbeitern (1992) die Verteilung GABAerger
Strukturen im Diencephalon von Hühnern mittels Licht- und
Elektronenmikroskopie. Ebenso wie zuvor von Domenici (1988) wurden im
Nucleus subpraetectalis vor allem die mittleren und großen GABAergen Somata
erkannt. Die Dendriten dieser Nervenzellen waren ebenfalls GABA-positiv.
Ausgehend vom Nucleus subpraetectalis stellten die Autoren außerdem
gebündelte Faserfragmente fest, die in den ventro-lateralen Nucleus rotundus
eintraten. Der Nucleus subpraetectalis und der Nucleus posteroventralis thalami
scheinen demnach die Quelle hemmender Einflüsse auf den Nucleus rotundus zu
sein.
Eine immunhistochemische und autoradiographische Untersuchung über die
Verteilung von GABAA-, GABAB- und Benzodiazepin-Rezeptoren zeigten im
Nucleus subpraetectalis nur Markierungen gegen den GABAA-Rezeptor, wobei
immunhistochemische Untersuchungen diese Ergebnisse bestätigen (Veenman et
al., 1994). Die Autoren bemerkten, daß Regionen mit hoher endogener GABA-
Konzentration eine geringe Rezeptormarkierung zeigen, vermutlich aufgrund der
Konkurrenz zwischen radioaktiv-markierter und endogener GABA.
Bei dem experimentellen Nachweis GABAerger Strukturen im Vorder- und
Mittelhirn der Taube wurde im Nucleus subpraetectalis und Nucleus interstitio-
praetecto-subpraetectalis eine dichte Ansammlung GAD- (Enzym zur GABA-
Synthese) und GABA-positiver Neuronen und eine leichte Markierung GAD-
positiver Fasern und Terminalien beobachtet (Veenman & Reiner, 1994). Die

Einleitung
22
Autoren vermuten auf Grund ihrer Daten verschiedene GABAerge Projektionen im
Taubenhirn, unter anderem auch die inhibitorische Projektion des Nucleus
subpraetectalis und des Nucleus interstitio-praetecto-subpraetectalis auf den
Nucleus rotundus.
Bei der Suche nach dem Ursprung eines vermutlich GABAergen Einflusses auf
den Nucleus rotundus führten Mpodozis und Mitarbeiter (1996)
immunhistochemische Untersuchungen sowie Tracing- und Läsionsexperimente
durch. Sie injizierten einen retrograden Tracer in den dorsoanterioren und
zentralen Nucleus rotundus, der zu starken ipsilateralen neuronalen
Markierungen, und bilateral zu axonalen Markierungen im Nucleus subpraetectalis
führte. Die Läsionsstudien wurden im Nucleus subpraetectalis durchgeführt. Der
Nucleus rotundus zeigte vor der Läsion des Nucleus subpraetectalis eine
GABAerge Anfärbung des Neuropil, wobei diese Markierung nach der
elektrolytischen Läsion des ipsilateralen Nucleus subpraetectalis deutlich abnahm.
Mpodozis und Mitarbeiter vermuteten daraufhin eine bilaterale tecto-rotundale
exzitatorische Innervation sowie eine indirekte ipsilaterale inhibitorische Projektion
vom Nucleus subpraetectalis, Nucleus interstitio-praetecto-subpraetectalis und
Nucleus posteroventralis auf den Nucleus rotundus.
1.2.4.4 Mögliche Funktionen der praetectalen Kerne
Auf Grund der Morphologie der Neuronen des Nucleus subpraetectalis der Hühner
postulieren Tömböl und Mitarbeiter (1999) diese als Interneuronen im tectofugalen
System. Die Autoren vermuten, daß es sich beim Nucleus subpraetectalis um eine
spezielle Neuronengruppe handelt, die modulatorische Eigenschaften auf Distanz,
daß heißt auf weitere Kerngebiete, zum Beispiel im tectofugalen System, ausübt.
Weitere Erkenntnisse über die funktionelle Relevanz der praetectalen Kerne
wurden in Verhaltensexperimenten vor und nach gezielter Läsion des Nucleus
rotundus, des Nucleus geniculatus pars ventralis und des Nucleus subpraetectalis
gewonnen (Hodos & Bonbright, 1975). Die Tiere wurden präoperativ auf die
Erkennung von Lichtunterschieden trainiert; die Läsionen wurden in drei Gruppen
unterteilt: die erste Gruppe erfuhr Läsionen im Nucleus rotundus, die zweite im
Nucleus rotundus und Nucleus geniculatis lateralis pars ventralis, und die dritte
Gruppe war durch Läsionen im Nucleus rotundus, Nucleus geniculatis lateralis
pars ventralis und Nucleus subpraetectalis gekennzeichnet. Im postoperativen

Einleitung
23
Training wurden vergleichbar hohe Defizite in der ersten und der dritten Gruppe,
daß heißt nach der Läsion des Nucleus rotundus beziehungsweise des Nucleus
rotundus, Nucleus geniculatis lateralis pars ventralis und Nucleus subpraetectalis
beobachtet. Gemeinsame Läsionen des Nucleus rotundus und des Nucleus
geniculatis lateralis pars ventralis führten hingegen zu geringen oder keinen
Veränderungen der Differenzierung von Helligkeitsunterschieden. Die Autoren
vermuten, daß zerstörte Zellgruppen, welche zu einer Beeinträchtigung der
visuellen Leistung führen, nicht zwangsweise visuelle Informationen verarbeiten.
Sie spekulieren dagegen, daß solche Zellgruppen modulierend auf die Aktivität
anderer Zellgruppen in einem parallelen System wirken könnten. Die Aufgaben
der indirekten tecto-praetecto-rotundalen Projektion könnten die Diskrimination
relevanter und irrelevanter visueller Stimuli, zum Beispiel bei der Futterauswahl,
sein (Veenman und Reiner, 1994b).
Zusammenfassend wird der Nucleus subpraetectalis als eine inhibitorisch über
GABAerge Mechanismen wirksame Zellgruppe angenommen (Domenici et al.,
1988; Granda & Crossland, 1989; Ngo et al., 1992; Veenman et al., 1994;
Veenman & Reiner, 1994; Mpodozis & et., 1996). Untersuchungen zu
Konnektivitäten im tectofugalen System zeigten, daß der SP/IPS-Komplex eine
kollaterale Verbindung zur Projektion des Tectum opticum zum Nucleus rotundus
darstellt (Benowitz & Karten, 1976; Hunt & Künzle, 1976a; Bischof & Niemann,
1990; Tömböl et al., 1994; Deng & Rogers, 1998). Eletrolytische Läsion des
Nucleus subpraetectalis führten zum Verschwinden GABAerger Projektion im
Nucleus rotundus. Eine direkte Wirkung des SP/IPS-Komplexes könnte demnach
eine zeitverzögerte Inhibition des tectorotundalen Informationsflusses sein.
1.3 Grundlagen der visuellen Systeme von Vögeln und Säugern
Vor etwa 300 Millionen Jahren existierten die Vorfahren der Reptilien, Vögel und
Säugetiere als „primitive“ Tetrapoden. Die Trennung der gemeinsamen
Entwicklung fand vor 280 Millionen Jahren statt. Im Laufe der Evolution
entwickelten sich die einzelnen Vertebratenklassen unabhängig voneinander,
gleichwohl auch heute noch einzelne Merkmale auf einen gemeinsamen Ursprung
zurückzuführen sind. Zum Beispiel können im Nervensystem zahlreiche
Gemeinsamkeiten zwischen Vögel, Reptilien und Säuger ausgemacht werden, wie

Einleitung
24
die Perzeption und Verarbeitung visueller Informationen in mehreren parallelen
Sehbahnen. Die im folgenden dargestellten Vergleiche im visuellen System von
Reptilien, Vögeln und Säugern beziehen sich auf eine Arbeit von Shimizu und
Karten (1993), die zahlreiche Hirnstrukturen im visuellen System dieser
Vertebratenklassen auf Analogien untersucht haben.
Eine der aufsteigenden Sehbahnen innerhalb dieser Vertebratenklassen verläuft
von der Retina zum Thalamus und von dort weiter zum Telencephalon. Bei Vögeln
und Reptilien wird sie als thalamofugale Sehbahn bezeichnet während das Analog
bei Mammaliern die retinogenikuläre Sehbahn ist. Die zweite, wichtige
aufsteigende Sehbahn bei Vögeln und Reptilien wird als tectofugale Sehbahn
bezeichnet. Sie verläuft von der Retina über das mesencephale Tectum opticum
zum diencephalen Nucleus rotundus, und terminiert schließlich im telencephalen
Ectostriatum. Bei Säugern wird die extragenikuläre Sehbahn als analog zu dieser
Sehbahn vermutet. Die extragenikuläre Sehbahn führt von der Retina über den
mesencephalen Colliculus superior zum thalamischen Pulvinar, und von dort
schließlich zum extrastriatalen Cortex.
Einzelne Kerne entlang dieser beiden aufsteigenden Sehbahnen, dem
tectofugalen System der Vögel und dem extragenikulären System der Säuger,
werden dabei als analog eingestuft. Zum Beispiel wird das Tectum opticum der
Vögel und Reptilien mit dem Superioren colliculus der Säuger, und der Nucleus
rotundus der Vögel und Reptilien mit dem thalamischen Pulvinar der Säuger
verglichen.
Das Telencephalon der Vögel, Reptilien und Säuger unterscheidet sich hingegen
wesentlich voneinander. Während bei Vögeln und Reptilien die Neuronen
hauptsächlich in Kerngebieten gruppiert sind, weist der Cortex der Säuger eine
charakteristische laminäre Strukturierung auf. Shimizu und Karten (1993) stellten
dennoch die Hypothese auf, daß Substrukturen des Ectostriatum der Vögel mit
distinkten Regionen des extrastriatären Cortex der Säuger verglichen werden
können, wie zum Beispiel der Area 19 und 21a. Ähnlichkeiten zwischen dem
Vorderhirn der Vögel und Reptilien sowie dem der Säuger zeigen sich in
Untersuchungen der Konnektivitäten und der biochemischen Eigenschaften. In
neurochemischen Untersuchungen wurden ähnliche Konstellationen von
Transmittern, Peptiden und Rezeptoren, wie zum Beispiel der Acetyltransferase,
der Glutamatdecarboxylase, Serotonin und GABAA-Rezeptoren im Telencephalon

Einleitung
25
der Vögel und der Säuger gefunden. Hingegen sind aber auch deutliche
Unterschiede in der Dichte neuroaktiver Substanzen auszumachen. Die
Ähnlichkeiten der Verteilung oben beschriebener Moleküle deutet auf eher geringe
Modifikationen hin, wodurch eine konservative Entwicklung und möglicherweise
auch Funktion im neuronalen System der Vögel und Säuger bekräftigt wird.
Unterschiede in der Verteilung von Transmittern, Peptiden und Rezeptoren bei
Vögeln und Säugern kennzeichnen vermutlich während der Evolution erfolgte
Veränderungen, möglicherweise Spezialisierung in dem jeweiligen System.
1.4 Neuronale Elemente der Signalübertragung
Mögliche exzitatorische und inhibitorische Signalwege zwischen dem Tectum
opticum, dem Nucleus subpraetectalis und dem Nucleus rotundus wurden in der
gegenwärtigen Arbeit mit Hilfe spezifischer Antikörper gegen diverse Glutamat-
Rezeptoren, den GABAA-Rezeptor sowie das GABA synthetisierende Enzym
Glutamatdecarboxylase (GAD) untersucht. Außerdem wurden
immunhistochemische Markierungen gegen Parvalbumin, einem Calcium-
bindenden Protein, in diesen Kerngebieten des visuellen Systems der Taube
durchgeführt. Im diesem Abschnitt werden die biochemischen Eigenschaften der
Signalübertragung auf neuronaler Ebene erläutert.
Neben der elektrischen Kopplung von Nervenzellen über gap-junction Proteine,
die einen schnellen Austausch von Ionen ermöglichen, kommunizieren Neuronen
durch die Ausschüttung von Transmittersubstanzen an ihren Nervenendigungen.
Nach Anbindung dieser an die entsprechenden Rezeptoren aktivieren oder
hemmen sie die elektrische Signalübertragung und schließlich die Funktion der
korrespondierenden Nervenzelle.
Ein Verständnis der Wirkungsweise dieser zwischen Neuronen ausgetauschten
Transmittersubstanzen erlaubt weitere Einblicke in die Mechanismen, die einer
entsprechenden Funktion dieser Neuronen und damit auch der
Verhaltenssteuerung zugrunde liegen.
Im Nervensystem sind bislang eine Vielzahl von Transmittern identifiziert worden,
wobei drei Gruppen von Substanzen unterschieden werden:
1. Aminosäurederivate (zum Beispiel Acetylcholin, Dopamin, Glutamat, ? -Aminobuttersäure (GABA))
2. Peptide (zum Beispiel Enkephaline, Vasopressin, Oxytocin)

Einleitung
26
3. Gase (zum Beispiel CO, NO)
In der vorliegenden Arbeit wurde die Verteilung und Wirkorte der beiden
klassischen Neurotransmitter Glutamat und GABA innerhalb der praetectalen
Kerne der Taube untersucht. L-Glutamat fungiert als exzitatorischer
Neurotransmitter im zentralen Nervensystem (Krnjevic, 1974), der seine Wirkung
über die Bindung an entsprechenden ionotrophen Glutamat-Rezeptoren (NMDA:
N-methyl-D-aspartat, Kainat; AMPA: ? -amino-3-hydroxy-5-methyl-isoxazole-4-
propriosäure) oder metabotrophen Glutamat-Rezeptoren ausübt (Watkins &
Evans, 1981; Fagg, 1985; Mayer & Westbrook, 1987b; Sladeczek et al., 1985).
GABA ist ein inhibitorischer Neurotransmitter des zentralen Nervensystems
(Krnjevic, 1974), der an den ionotrophen GABAA-Rezeptor oder den
metabotrophen GABAB-Rezeptor binden kann (Dunlap, 1981; Hill & Bowery, 1981;
Bormann, 1988; Nicoll, 1988). Ionotrophe und metabotrophe Rezeptoren
unterscheiden sich unter anderem dahingehend, daß bei den ionotrophen
Rezeptoren ein für Ionen permeabler Kanal direkter Bestandteil des Rezeptors ist,
wobei die Bindung des Transmitters eine Konformationsänderung des Ionenkanals
bewirkt (Eccles & McGeer, 1979). Die Rezeptoraktivierung wirkt sich somit direkt
auf die Permeabilität des Ionenkanals aus. Bei den metabotrophen Rezeptoren
hingegen bewirkt die Transmitterbindung die Aktivierung einer „second-
messanger“ Kaskade über ein Guanin Nucleotid-Bindungsprotein (G-Protein), als
deren Folge ein räumlich benachbarter Ionenkanal in seinem Öffnungsverhalten
beeinflußt wird (Eccles & McGeer, 1979).
1.4.1 Ionotrophe Glutamat-Rezeptoren
In der klassischen Nomenklatur werden die ionotrophen Glutamat Rezeptoren
nach ihrer Aktivierbarkeit durch den Agonisten NMDA in non-NMDA Rezeptoren
und NMDA Rezeptoren unterteilt. Die non-NMDA Rezeptoren wiederum werden
aufgrund ihrer bevorzugten Aktivierung durch die Agonisten AMPA und Kainat
untergliedert. Gemeinsames strukturelles Motiv der Glutamat-Rezeptoren ist der
Aufbau aus jeweils fünf Rezeptoruntereinheiten, wobei die durch AMPA
aktivierbaren Glutamat-Rezeptoruntereinheiten Glu R1, Glu R2, Glu R3 und Glu
R4 von den durch Kainat stimulierbaren Rezeptoruntereinheiten Glu R5, Glu R6,
Glu R7, KA-1 und KA-2 unterschieden werden. Von diesen abgrenzbar sind
wiederum die NMDA Rezeptoruntereinheiten NR1, NR2A, NR2B, NR2C und

Einleitung
27
NR2D. Die verschiedenen Rezeptoruntereinheiten entstehen durch alternatives
Schneiden und Prozessieren des Glutamatrezeptorgens. Der Aufbau des
Rezeptors aus den entsprechenden AMPA, Kainat oder NMDA
Rezeptoruntereinheiten bestimmt schließlich die charakteristischen
Öffnungseigenschaften des integrierten Ionenkanals. Während AMPA und Kainat
Rezeptoren nach der physiologischen Bindung von Glutamat zumeist permeabel
für nur einwertige Ionen wie Natrium sind, so können nach massiver Erregung der
postsynaptischen Membran durch Glutamat, zweiwertige Ionen wie Calcium den
NMDA Rezeptorionenkanal passieren. AMPA Rezeptoren, bei denen die
Rezeptoruntereinheit Glu R2 nicht Bestandteil des Rezeptors ist, sind darüber
hinaus auch permeabel für Calcium. Da Calcium in Nervenzellen als sogenannter
intrazellulärer zweiter Botenstoff vielfältige Signalkaskaden beeinflußt, wirkt ein
Glutamat-Rezeptor vermittelter postsynaptischer Calcium Einstrom stark regulativ
auf das Signal empfangende Neuron.
1.4.2 Ionotrophe GABA-Rezeptoren
Der im zentralen Nervensystem am weitesten verbreitete ionotrophe GABA-
Rezeptor ist der GABAA-Rezeptor. Sein struktureller Aufbau ist vermutlich ähnlich
dem der ionotrophen Rezeptoren allgemein, daß heißt fünf
Rezeptoruntereinheiten formen sich zu einem funktionsfähigen Rezeptor
zusammen. Bislang sind acht verschiedene Rezeptoruntereinheiten bekannt (? , ? ,
? , ? , ? ,? ,? und ? ) (Whiting et al., 1995; Sieghart et al., 1995; Hedblom & Kirkness,
1997; Whiting et al., 1997; Whiting, 1999). Während die Untereinheiten ? , ? , ? und
? ubiquitär um Gehirn verbreitet sind, so ist die Expression von ? auf die Retina
beschränkt. Die Rezeptoruntereinheiten ? ,? und ? wurden erst in jüngerer Zeit
charakterisiert, und ihr Vorkommen ist vermutlich vornehmlich auf das periphere
Nervengewebe begrenzt (Hedblom & Kirkness, 1997; Whiting et al., 1997;
Whiting, 1999).
Die verschiedenen Rezeptoruntereinheiten sind in diversen Kombinationen von
Heteromeren zu GABAA-Rezeptoren zusammengebaut, wodurch sich die
elektrophysiologischen und pharmakologischen Eigenschaften ergeben.
Grundsätzlich ist der GABAA-Rezeptor selektiv permeabel für Anionen wie Chlorid,
wodurch es nach Bindung von GABA im Nervensystem der Vertebraten zu einer

Einleitung
28
Hyperpolarisation der Zelle durch einen Chlorid-Ionen-Einstrom kommt.
Intrazellulären Einfluß auf den GABA-Rezeptor haben die Proteinkinasen A und C
sowie Calcium-Calmodulin abhängige Proteinkinasen.
1.4.3 Glutamatdecarboxylase
Das für die Synthese des Neurotransmitters GABA wichtige Enzym ist die
Glutamatdecarboxylase (GAD), da es die Umwandlung von Glutamat in GABA
katalysiert. Seine Funktion bei der Herstellung von GABA und sein spezifisches
zytosolisches Vorkommen machen es zu einem idealen Marker für GABAerge
Neuronen im Nervensystem.
1.4.4 Parvalbumin
Parvalbumin (PV) gehört zu den Calcium-bindenden Proteinen, die im
Nervensystem der Vertebraten weit verbreitet sind (Celio & Heizman, 1981; Celio
et al., 1990). PV fungiert allgemein als regulatives Protein, welches die Calcium-
Homöostase von Nervenzellen gewährleistet (Heizmann, 1984; Celio, 1990).
Häufig ist PV in GABAergen, schnell feuernden Neuronen, lokalisiert (Celio, 1986;
Freund, 1989).
Eine physiologische Bindung von Calcium-Ionen in Zellen wird notwendig, da
Calcium-Ionen in diesen die Funktionen intrazellulärer, zweiter Botenstoffe
erfüllen, und an einer Vielzahl von Prozessen in Neuronen beteiligt sind. Über die
Regulation der Phosphorylierung-Dephosphorylierung verschiedener Proteine
steuern sie zum Beispiel Prozesse der Transmitterfreisetzung (Smith & Augustine,
1988), oder mikrotubuläre Transportprozesse (Chan et al., 1980), ebenso wie die
Funktionszustände von Rezeptoren (Greengard et al., 1996).

Einleitung
29
1.5 Zielsetzung der Arbeit
In der vorliegenden Arbeit wurde die Integration des Nucleus subpraetectalis /
Nucleus interstitio-praetecto-subpraetectalis (SP/IPS) innerhalb der tectofugalen
Informationsverarbeitung untersucht. Vorausgegangene Studien deuteten an, daß
das SP/IPS System vermutlich inhibitorischen, GABAergen Einfluß auf das
tectofugale System nimmt. Injektionen neuronaler Farbstoffe in den Nucleus
rotundus (RT) gaben auf Grund retrograder Markierungen im SP/IPS Anzeichen
für eine Projektion dieser praetectalen Kerne auf den RT.
? ? Konnektivitäten des SP/IPS-Systems zum tectofugalen System
Durch Injektionen anterograd transportierter neuronaler Marker wie Biotin-
Dextran-Amin und die ß-Untereinheit des Choleratoxin in den SP/IPS sollte
eine Verbindungen zum RT verifiziert werden. Darüber hinaus sollten durch den
ebenfalls gegebenen retrograden Transport dieser neuronalen Marker der
afferente Input der praetectalen Kerne (SP/IPS) von Neuronen der tectalen
Schicht 13 verifiziert werden.
? ? Charakterisierung biochemische Signalwege im SP/IPS-Komplex
Mit Hilfe verschiedener Antikörper sollten mögliche exzitatorische (glutamaterge)
und inhibitorische (GABAerge) Signalwege im SP/IPS-Komplex erfaßt werden.
Außerdem wurde die Expression des Calcium-bindenden Proteins Parvalbumin in
diesen praetectalen Kernen erfaßt. Quantitative Analysen der immunpositiven
Neuronen sollten Aufschluß über mögliche Heterogenitäten der
Transmitterverteilung zwischen beiden Hemisphären, der anterior-posterior
Ausdehnung, sowie zwischen dem SP und dem IPS geben.
? ? Einfluß des SP/IPS-Komplexes auf das tectofugale System
Anhand der unter Punkt 1 gewonnen Daten zur Konnektivität des SP/IPS-Systems
zum tectofugalen System, sowie der unter Punkt 2 erlangten Daten über mögliche
exzitatorische und inhibitorische Signalwege innerhalb des SP/IPS-Systems, sollte
ein Modell zum Einfluß dieser praetectalen Kerne auf die Informationsverarbeitung
im tectofugalen System erarbeitet werden.

Material und Methoden
30
2 Material und Methoden
2.1 Material
Die Tracing-Untersuchungen und immunhistochemischen Studien wurden im
Nucleus subpraetectalis der Taube Columba livia durchgeführt. Für die Tracing-
Untersuchungen standen insgesamt 12 Tiere, für die immunhistochemischen
Untersuchungen insgesamt 9 Tiere beiden Geschlechts, im Alter von 1 bis 10
Jahren zur Verfügung. Sie wurden aus der lokalen Zucht bezogen; die
durchgeführten Untersuchungen wurden gemäß der deutschen Gesetzgebung des
Tierschutzes durchgeführt.
2.2 Tracing
Bei der Tracing-Technik werden mittels einer Stereotaxie Farbstoff-Injektionen in
definierte Hirnareale durchgeführt, um Verbindungen verschiedener Kerngebiete
zu erfassen. Durch Zusatz von Detergenzien (DMSO bei BDA) wird hierbei eine
erhöhte Membranpermeabilität hervorgerufen, wodurch die injizierten Substanzen,
die auch als Tracer bezeichnet werden, in die Zelle eindringen können. Tracer
werden entlang der Fortsätze von Neuronen transportiert und erlauben somit die
Darstellung des dentritischen und axonalen Verzweigungsmusters. Die genaue
Lokalisation des Injektionsortes, in der gegenwärtigen Arbeit der mesencephale
Nucleus subpraetectalis der Taube, wurde anhand des „Stereotaxic Atlas of the
Brain of the pigeon (Columbia livia)“ (Karten & Hodos, 1967) ermittelt. Für die
Tracingexperimente standen insgesamt zwölf Tiere mit einem Gewicht von 464-
570g zur Verfügung.
2.3 Tracer
In der vorliegenden Arbeit wurden als Tracer Biotin-Dextran-Amin (BDA) sowie die
? -Untereinheit des Choleratoxins (CtB) verwendet. Der Tracer BDA wurde
neunmal injiziert, und CtB wurde sechsmal verwendet. Bei fünf der zwölf Tiere
wurden bilaterale Injektionen durchgeführt, wobei jeweils rechts oder links BDA,
beziehungsweise CtB injiziert wurden.

Material und Methoden
31
2.3.1 BDA
Bei BDA handelt es sich um einen bevorzugt anterograd transportierten Tracer mit
einem Molekulargewicht von 10.000 MW (Molecular Probes, Holland). Das BDA
wurde als 10%-ige Lösung in 4% DMSO verwendet. Um eine ausreichende
Markierung zu erreichen, wurde in jede Taube ein Tracervolumen von 36 bis 75nl
durch Druckinjektionen (WPI-Nanoliter-Injektor, Harry Fein, Berlin) appliziert.
2.3.2 CtB
CtB ist ein Tracer, der sowohl retro-, als auch anterograd transportiert wird. Das
Tracervolumen lag bei zwischen 23 und 75nl im Nucleus subpraetectalis.
2.4 Tracingverfahren
Die Tiere wurden zunächst mit Equitesin (0,33ml/100g KG) narkotisiert, und
anschließend auf einem Wärmekissen in eine stereotaktische Apparatur eingelegt.
Der antero-posteriore Nullpunkt der Taube wurde hier durch Fixierung der
Gehörgänge und des Schnabels festgelegt. Der benötigte Tracer, BDA
beziehungsweise CtB, wurde vorsichtig in eine Glaspipette aufgesogen, die mit
einem Druckinjektor verbunden war. Nach einem kleinem Schnitt in die Kopfhaut
wurde der medio-laterale Nullpunkt des Schädels mittels eines Loches in der
Schädeldecke und der Lokalisation des Sinus sagittalis superior, bestimmt.
Anhand des Taubenatlas nach Benowitz & Karten (1967) wurde die Stereotaxie in
der gewünschten antero-posterioren und medio-lateralen Ebene eingestellt, und
an der entsprechenden Stelle vorsichtig ein kleines Loch in die Schädeldecke
gebohrt. Nachfolgend wurde die Glaspipette auf die Gehirnoberfläche aufgesetzt,
und die Stereotaxie entsprechend in der dorso-ventralen Achse geeicht. Nach
Berechnung der Zielkoordinate in der dorso-ventral Ebene wurde die Glaspipette
bis zur gewünschten Eindringtiefe vorgeschoben. Nach Erreichen des
Injektionsortes wurde zur Adaptation des umgebenden Gewebes an den
Fremdkörper ca. 10 Minuten gewartet. Die folgende Injektion verlief langsam über
einen Zeitraum von 5-10 Minuten, wobei abschließend die Glaspipette für 10
Minuten am Injektionsort belassen wurde, um die Verteilung des Tracers im
Injektionskanal zu vermeiden.

Material und Methoden
32
2.5 Überlebenszeiten
Die Laufeigenschaften des Tracers bestimmen den Zeitraum von der Injektion bis
zur Perfusion der Tiere. Bei BDA wurde drei bis sieben Tage, bei CtB drei bis fünf
Tage gewartet, bevor wie unter Punkt 2.6 beschrieben perfundiert wurde.
2.6 Perfusion
Zur Fixierung der Hirnstrukturen der Taube wurde das Tier zunächst mit einer
Kochsalzlösung (0.9% NaCl) und anschließend mit einem entsprechenden Fixativ,
bestehend aus 4% Paraformaldehyd + 0,1% Glutaraldehyd in 0,12 M
Phosphatpuffer, perfundiert. Vor Beginn der Perfusion wurde der Taube 0,2ml
Heparin (1000Units) intramuskulär in den Brustmuskel verabreicht. Das Heparin
vermeidet eine vorzeitige Blutgerinnung, welche zu einer Verstopfung des
Gefäßsystems führen könnte. Zehn Minuten später erfolgte die Anästhesie mit
Equitesin (0,5ml/100g Körpergewicht (KG)), erneut intramuskulär in den
Brustmuskel. Sobald sich das Tier in tiefer Narkose befand, wurden die Federn
des Thorax gerupft. Die Eröffnung des Brustkorbes begann mit einem Schnitt
entlang der unteren Rippenbögen, wobei die Leber durch vorsichtiges Ausschälen
der inneren Thoraxwand gelöst wurde. Nach der Eröffnung des Perikards erfolgte
das Einführen des Katheters in den linken Herzventrikel, ein kleiner Schnitt
oberhalb des rechten Vorhofes ermöglichte den Abfluß des Blutes aus dem
Kreislaufsystem, und die Perfusion mit 40?C warmer 0.9% NaCl Lösung wurde
begonnen. Sobald die Skleren der Augen blutleer erschienen, meist nach der
Perfusion von ca. 200ml NaCl, wurde mit der Fixierlösung perfundiert. Das
Zeichen einer guten Perfusion war eine Versteifung und Überstreckung des
Kopfes.
2.7 Postfixieren
Das Gehirn wurde vorsichtig aus dem Kopf herauspräpariert, und anschließend für
zwei Stunden postfixiert. Die Zusammensetzung der Postfixierlösung bestand aus
der bereits bei der Perfusion verwendeten Fixierlösung, in welcher zusätzlich 30%
Succrose gelöst wurden.

Material und Methoden
33
2.8 Schneiden der Gehirne
Für die weiteren Untersuchungen wurden 35µm dicke Frontalschnitte des Gehirns
auf einem Kryostaten angefertigt. Als Einbettmedium wurde „Tissue Freezing
Medium“ (Jung, Deutschland) benutzt. Die Schneidetemperatur lag bei minus
25?C. Die Hirnschnitte wurden dann in einer PBS-Lösung mit 0,1% Natriumazid
(NaN3) gesammelt. NaN3 hatte die Aufgabe die Pilz- und Bakterienbildung zu
unterdrücken.
2.9 Darstellung von BDA
Um die Markierungen von BDA zu visualisieren, wurde ein Vectastain ABC Elite
Kit (Vector Laboratories, Camon, Wiesbaden) verwendet. Zuvor wurden die
Schnitte zur Ausschaltung endogener Peroxidasen für 30 Minuten in 0.1 % H2O2
inkubiert. Danach wurden die Gehirnschnitte 3 mal 2 Minuten und 2 mal 10
Minuten in 0,12 M PBS gespült, bevor die unter Punkt 2.18 beschriebene ABC-
Reaktion folgte. Nach der ABC-Reaktion wurden die Schnitte erneut drei mal zwei
Minuten in 0,12 M PBS und zwei mal zehn Minuten in 0,1M Na-Acetatpuffer (pH
6,0) gespült. Anschließend folgte die Visualisierung des Tracers an welchen der
ABC-Komplex, und somit die Peroxidasen gebunden hatte, mittels der DAB-
Reaktion (Punkt 2.19).
2.10 Darstellung von CtB
Die Darstellung von CtB erfolgte durch eine immunhistochemische Behandlung
der Gehirnschnitte. Begonnen wurde hierbei mit einer Inkubation der Hirnschnitte
für 30 Minuten in 0,1 % H2O2 zur Inaktivierung endogener Peroxidasen. Danach
wurden die Hirnschnitte 3 mal 2 Minuten und 2 mal 10 Minuten in 0,12 M PBS
gespült. Es folgte eine Inkubation der Proben in Normalserum (Affenserum,
Jackson, USA) in 10ml PBS und 0,1ml Triton X 100 (PBS-T) für 30 Minuten bei
Raumtemperatur. Das Serum stammte vom Spender des sekundären Antikörpers
und diente dazu, unspezifische Anbindungsstellen für die Antikörper zu blockieren.
Nach erneutem Spülen wurde über Nacht (4°C) im primären Antikörper, anti-CtB
(1:10000 in PBS-T), inkubiert. Am nächsten Tag wurde kurz mit PBS gespült,
bevor die Hirnschnitte für 60 Minuten bei Raumtemperatur im sekundärem

Material und Methoden
34
Antikörper inkubierten. Anschließend folgte eine ABC- und darauf eine DAB-
Reaktion (Punkt 2.18 und 2.19).
Bei den sechs bilateral BDA und CtB injizierten Tauben, wurde die histologische
Gewebeaufbereitung verändert, um eine Differenzierung der beiden Tracer zu
ermöglichen. Zur Darstellung von BDA wurden die Hirnschnitte zunächst einer
ABC- Reaktion und einer anschließenden Kobalt-Nickel-intensivierten DAB-
Reaktion unterzogen (Punkt 2.18 / 2.19), worauf die beschriebene
immunhistochemische Detektion mit anti-CtB erfolgte. Bei der abschließenden
DAB-Reaktion wurde zur Darstellung von CtB auf eine Nickel-Kobalt
Intensivierung verzichtet. Das so erhaltene Signal war heller und rötlicher als das
der BDA-Markierungen.
2.11 Auswertung
Die Auswertungen der Tracing-Untersuchungen erfolgten an einem Mikroskop der
Firma Olympus, Typ BH 2, wobei die mikroskopischen Bilder im Rechner
digitalisiert wurden, und die markierten Zellstrukturen anschließend mit Hilfe eines
Bildanalysesystems („analysis“, SIS, Münster, Deutschland; „designer“,
Micrografx, Dallas, USA) nachgezeichnet werden konnten.
2.12 Immunhistochemie
Bei der Methode der Immunhistochemie (IHC) werden Proteine, zum Beispiel
Enzyme, Transmitter oder Rezeptoren, durch spezifische Antikörper markiert. Die
Prozedur im einzelnen sowie die ablaufenden Reaktionen werden in den
nachfolgenden Abschnitten dargestellt. Ein wichtiger Faktor für eine gelungene
Darstellung von Antigenen mittels der IHC ist eine geeignete Vorbehandlung und
Fixierung des zu untersuchenden Gewebes sowie der Einsatz spezifischer
Antikörper.
2.13 Allgemeine Antikörperherstellung
Antikörper gehören zu den Gammaglobulinen und stellen eine heterogene Gruppe
von Glykoproteinen dar. Sie werden durch B-Lymphozyten und Plasmazellen bei
Kontakt des Organismus mit Antigenen gebildet (Antigen-Antikörper-Reaktion).

Material und Methoden
35
Spezifische Antikörper stellt man in Säugetieren wie Maus, Kaninchen oder Ziege
her. Zuerst wird dabei das in späteren Versuchen zu detektierende Protein, z.B.
eine gereinigte Rezeptoruntereinheit, subkutan injiziert. Dieses Protein löst nun als
Antigen eine Immunantwort aus, mit der Folge einer spezifischen
Antikörperproduktion gegen das Antigen. In der Phase der höchsten
Antikörperproduktion des Tieres entnimmt man Blut, und zentrifugiert dieses,
wodurch die Erythrozyten von dem restlichen antikörperreichen Serum getrennt
werden. Im Serum enthalten sind sogenannte polyklonale Antikörper, das heißt
außer den spezifisch gegen das Antigen gerichteten Antikörpern auch weitere
Antikörper, welche das Tier im Laufe seines Lebens gegen andere Antigene
gebildet hat.
Bei kleinen Antigenen kann durch eine Vergrößerung des Proteins, z.B. durch die
Anlagerung eines Aldehydes, eine verbesserte Immunantwort hervorgerufen
werden. Ein solches Verfahren erfordert häufig eine angepaßte Vorbehandlung
des zu untersuchenden Gewebes, wie zum Beispiel eine entsprechende
Fixierung. Die käuflich erwerbbaren Antikörper sind meist gereinigte Substanzen,
die eine optimale Immunreaktion zulassen.
2.14 Eingesetzte Antikörper
Die in der gegenwärtigen Untersuchung verwendeten primären Antikörper (pAK)
richten sich gegen den Neurotransmitter ? -Aminobuttersäure (GABA), das GABA
synthetisierende Enzym Glutamatdecarboxylase (GAD), gegen die ? -
Rezeptoruntereinheit des GABAA- Rezeptors, gegen das Calcium-bindende
Protein Parvalbumin (PV) und gegen die ionotrophen Glutamatrezeptor-
Untereinheiten vom AMPA–Typ (GluR2/3 und GluR4). Die sekundären Antikörper
(sAK) binden spezifisch an den pAK (Überblick: Tabelle 1).
2.14.1 Anti-Glutamatdecarboxylase
Zur Bildung des Antigens für die Antikörperproduktion gegen
Glutamatdecarboxylase (GAD) wurde geklonte DNA verwendet, die in E.coli
exprimiert wurde. Der polyklonale Antikörper richtet sich gegen die lange Sequenz
der GAD (~67kD; Kaufmann et al., 1986). Die Antikörper sind erhältlich bei
„Chemicon International INC“ unter der Katalognummer AB 108.

Material und Methoden
36
2.14.2 Anti-GABAA-Rezeptor (? -Untereinheit)
Die monoklonalen Antikörper gegen die ? -Untereinheit des GABAA-Rezeptor
stammen aus der Maus. Zu beziehen ist der anti-GABAA-Rezeptor Antikörper bei
der Boehringer Mannheim GmbH, Deutschland.
(Katalognummer: 1381458)
Literatur: Schoch & Möhler (1983), Möhler et al. (1986), Richards et al. (1987),
Watson et al. (1986).
2.14.3 Anti-AMPA-Rezeptoren
Die polyklonalen Antikörper richten sich gegen synthetisierte C-terminale
Aminosäure-Sequenzen der AMPA-Rezeptor-Untereinheiten der Ratte, und
wurden im Kaninchen hergestellt.
Glu R2/3 EGYNVYGIESVKI = 13 Aminosäuren (AS)
Glu R4 RQSSGLAVIASDLP = 14 AS
Die Oligopeptide wurden für 90 min bei Raumtemperatur mit 0,1% Glutaraldehyd
in einer phosphat-gepufferten Salzlösung (PBS) und Rinderserum-Albumin
konjugiert, bevor sie einem Kaninchen injiziert wurden. Der Vertrieb dieser
Antikörper erfolgt durch „Chemicon International INC“
(Katalognummer: AB 1506 (Glu R2/3); AB 1508 (Glu R4)).
Literatur: Petralia & Wenthold, 1992; Wenthold, 1992; Keinänen et al., 1990;
Wenthold, 1990.
2.14.4 Anti-Parvalbumin
Für die Herstellung des Antikörpers gewinnt man PV als Antigen aus der
Muskulatur von Karpfen. Der in Mäusen hergestellte monoklonale Antikörper
erkennt das Protein PV in den Nervenzellen verschiedener Säuger und Vögeln.
Erhältlich ist der Antikörper bei „Sigma“ unter der Katalognummer PA-235.
Literatur: Celio et al., 1990.
2.15 Sekundäre Antikörper
Zur Herstellung von Dauerpräparaten mittels der DAB-Reaktion wurden je nach
pAK polyklonale affinitätsgereinigte sAK eingesetzt. Der sAK gegen die pAK GAD,
Material und Methoden
37
Glu R2/3 und Glu R4 stammte aus einem ABC-Elite-Kit (Vector Laboratories,
Camon Deutschland). Die Spezifität dieses sAK ist Ziege gegen Kaninchen. Der
von Chemicon vertriebene sAK aus Kaninchen richtete sich gegen
Immunglobuline der Maus (Katalognummer: AP 160B), und wurde zur Detektion
des pAK gegen den GABAA-Rezeptor eingesetzt. Ein weiterer sAK aus einem
ABC-Elite-Kit war Ziege-anti-Maus, und wurde zur Detektion des pAK gegen PV
eingesetzt.
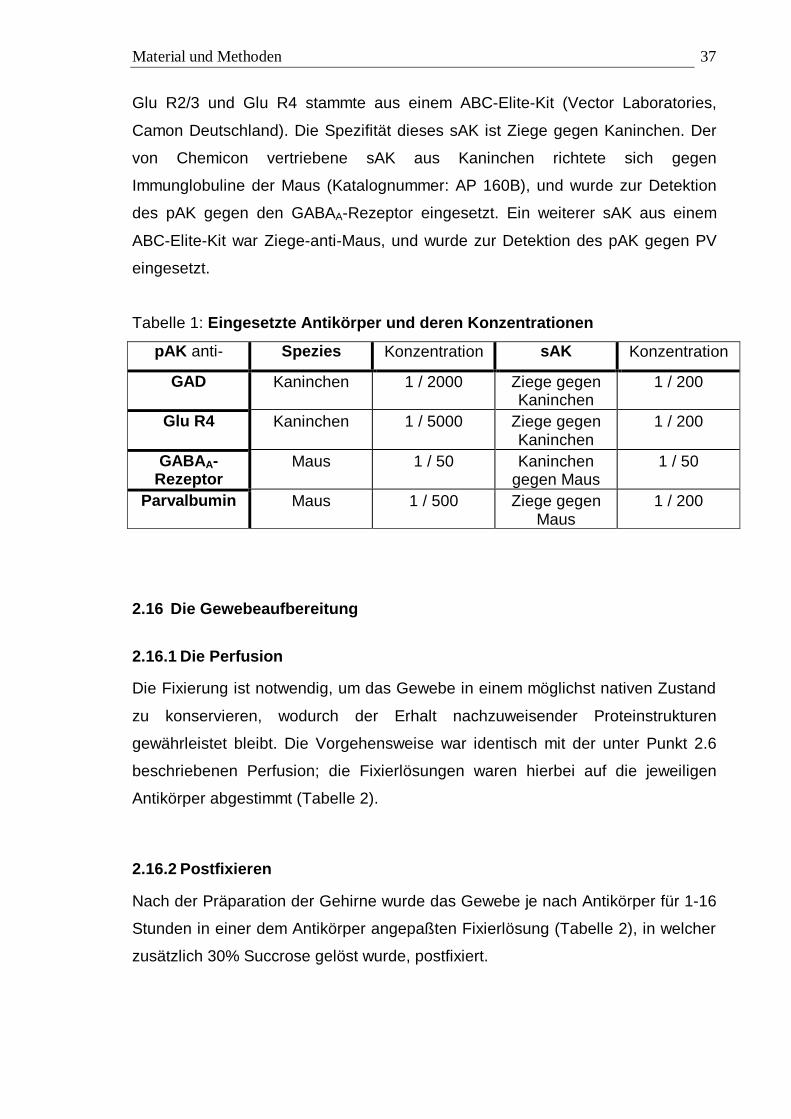
Tabelle 1: Eingesetzte Antikörper und deren Konzentrationen
pAK anti- Spezies Konzentration sAK Konzentration
GAD Kaninchen 1 / 2000 Ziege gegen Kaninchen
1 / 200
Glu R4 Kaninchen 1 / 5000 Ziege gegen Kaninchen
1 / 200
GABAA-Rezeptor
Maus 1 / 50 Kaninchen gegen Maus
1 / 50
Parvalbumin Maus 1 / 500 Ziege gegen Maus
1 / 200
2.16 Die Gewebeaufbereitung
2.16.1 Die Perfusion
Die Fixierung ist notwendig, um das Gewebe in einem möglichst nativen Zustand
zu konservieren, wodurch der Erhalt nachzuweisender Proteinstrukturen
gewährleistet bleibt. Die Vorgehensweise war identisch mit der unter Punkt 2.6
beschriebenen Perfusion; die Fixierlösungen waren hierbei auf die jeweiligen
Antikörper abgestimmt (Tabelle 2).
2.16.2 Postfixieren
Nach der Präparation der Gehirne wurde das Gewebe je nach Antikörper für 1-16
Stunden in einer dem Antikörper angepaßten Fixierlösung (Tabelle 2), in welcher
zusätzlich 30% Succrose gelöst wurde, postfixiert.
Material und Methoden
38
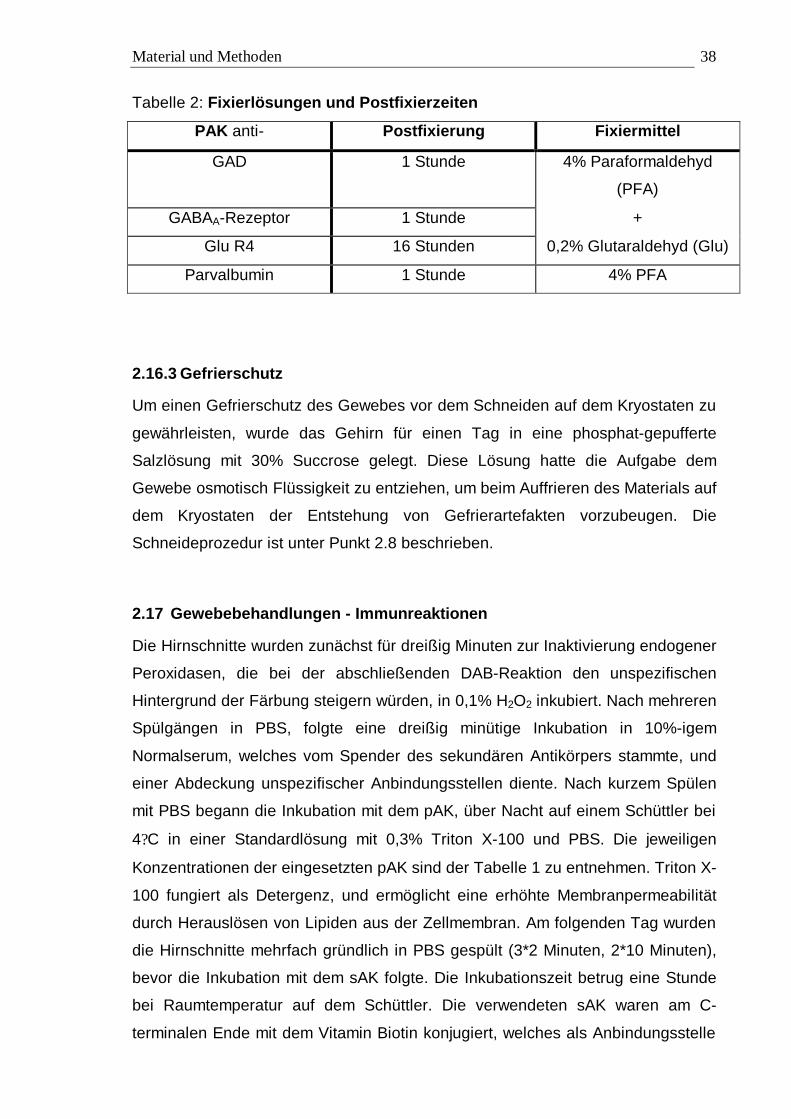
Tabelle 2: Fixierlösungen und Postfixierzeiten
PAK anti- Postfixierung Fixiermittel
GAD 1 Stunde 4% Paraformaldehyd
(PFA)
GABAA-Rezeptor 1 Stunde +
Glu R4 16 Stunden 0,2% Glutaraldehyd (Glu)
Parvalbumin 1 Stunde 4% PFA
2.16.3 Gefrierschutz
Um einen Gefrierschutz des Gewebes vor dem Schneiden auf dem Kryostaten zu
gewährleisten, wurde das Gehirn für einen Tag in eine phosphat-gepufferte
Salzlösung mit 30% Succrose gelegt. Diese Lösung hatte die Aufgabe dem
Gewebe osmotisch Flüssigkeit zu entziehen, um beim Auffrieren des Materials auf
dem Kryostaten der Entstehung von Gefrierartefakten vorzubeugen. Die
Schneideprozedur ist unter Punkt 2.8 beschrieben.
2.17 Gewebebehandlungen - Immunreaktionen
Die Hirnschnitte wurden zunächst für dreißig Minuten zur Inaktivierung endogener
Peroxidasen, die bei der abschließenden DAB-Reaktion den unspezifischen
Hintergrund der Färbung steigern würden, in 0,1% H2O2 inkubiert. Nach mehreren
Spülgängen in PBS, folgte eine dreißig minütige Inkubation in 10%-igem
Normalserum, welches vom Spender des sekundären Antikörpers stammte, und
einer Abdeckung unspezifischer Anbindungsstellen diente. Nach kurzem Spülen
mit PBS begann die Inkubation mit dem pAK, über Nacht auf einem Schüttler bei
4?C in einer Standardlösung mit 0,3% Triton X-100 und PBS. Die jeweiligen
Konzentrationen der eingesetzten pAK sind der Tabelle 1 zu entnehmen. Triton X-
100 fungiert als Detergenz, und ermöglicht eine erhöhte Membranpermeabilität
durch Herauslösen von Lipiden aus der Zellmembran. Am folgenden Tag wurden
die Hirnschnitte mehrfach gründlich in PBS gespült (3*2 Minuten, 2*10 Minuten),
bevor die Inkubation mit dem sAK folgte. Die Inkubationszeit betrug eine Stunde
bei Raumtemperatur auf dem Schüttler. Die verwendeten sAK waren am C-
terminalen Ende mit dem Vitamin Biotin konjugiert, welches als Anbindungsstelle

Material und Methoden
39
für die weitere ABC-Reaktion diente. Vor der ABC-Reaktion wurden die
Gehirnschnitte erneut intensiv mit PBS gespült (3*2 Minuten, 2*10 Minuten).
2.18 ABC-Reaktion
Über die ABC-Reaktion wurde die IHC-Reaktionskaskade mit einer Peroxidase
verbunden, welches in einer abschließenden Diaminobenzidin-Reaktion ein
Chromogen bindet und färbt. ABC steht für den Avidin-Biotin-Komplex, wobei das
Emzym Meeretichperoxidase an das Biotin gekoppelt war („ABC-Elite-Kit“, Vector
Laboratories, England). Zunächst wurden die Stammlösungen mit den einzelnen
Reaktionskomponenten gemäß Herstellerangaben dreißig Minuten vor Beginn der
ABC-Reaktion zusammengefügt. In einer neunzig minütigen Inkubation der
Hirnschnitte in der ABC-Lösung bei Raumtemperatur auf dem Schüttler, fand
anschließend eine Kopplung über das Avidin am biotinylierten sAK statt. Als letzter
Schritt dieser Reaktion wurde das Gewebe drei mal zwei, und zwei mal zehn
Minuten in PBS gespült.
2.19 DAB-Reaktion
Für die Detektion des Antigens nach immunhistochemischer Behandlung wurde
die langsame, aber gleichmäßig ablaufende Nickel-Kobalt intensivierte
Diaminobenzidin-Reaktion (DAB-Reaktion) nach Hsu und Soban (1982) und
Hancock (1986) durchgeführt. Hierbei wurde mittels eines Oxidationsprozesses
das primär farblose 3,3-Diaminobenzidin in eine braune Form überführt, die
mikroskopisch sichtbar wurde.
Zunächst wurde das Material zur Ansäuerung in Natrium-Acetat (pH 6.0) gespült,
um eine gute Farbintensität durch das Diaminobenzidin zu erreichen.
Anschließend folgte eine fünfzehn minütige Inkubation in einem Cocktail
bestehend aus Ammonium-Nickelsulfat, Ammonium-Chlorid, Kobaltchlorid und ß-
D-Glucose. Ammonium-Nickelsulfat, Ammonium-Chlorid und Kobaltchlorid
bewirkten eine Signalverstärkung. Im nächsten Schritt wurde einer Lösung
gleicher Zusammensetzung Glucoseoxidase (10µl, Stamm: 0,001g/ml) zugefügt.
Die Glucoseoxidase spaltete die ? -D-Glucose, setzt dabei kontinuierlich H2O2 frei,
und führt durch Reaktion mit den Peroxidasen zur Oxidation des DAB. Die

Material und Methoden
40
Reaktionsdauer betrug etwa 15 Minuten, wonach die Hirnschnitte zunächst in Na-
Acetat und dann in PBS gespült wurden.
2.20 Kontrollen
Zur Überprüfung der Spezifität der pAK und sAK wurden Kontrollexperimente
durchgeführt, bei denen der pAK durch ein Serum vom Spender des
Primärantikörpers ersetzt wurde, wobei der weitere Verlauf der IHC unverändert
blieb. Desweiteren wurde zur Überprüfung der Spezifität der sAK eine IHC ohne
pAK-Inkubation durchgeführt.
2.21 Färbung mit Kresylviolett
Zur allgemeinen Darstellung der mikroskopischen Morphologie des Taubenhirns
wurden Kresylviolett-Färbungen durchgeführt. Der Farbstoff Kresylviolett markierte
unabhängig vom Zelltyp das endoplasmatische Reticulum der Zellen. Dazu
wurden die Hirnschnitte auf gelatinisierten Objektträgern aufgezogen und
getrocknet. Es folgte ein kurzes Spülen in Aqua dest, gefolgt von einem Spülen in
70% Alkohol, zur Permeabilisierung des Gewebes, bevor das Material in mit
Essigsäure (99%) versetztem Aqua dest. (5 Tropfen Essigsäure pro 100ml Aqua
dest.) angesäuert wurde. Die Farbreaktion erfolgte für zehn Minuten in einer 1%-
igen Kresylviolett-Lösung. Danach wurden die Hirnschnitte kurz in Aqua dest.
gespült; eine weitere Differenzierung der Färbung fand in 70% und 96% Alkohol,
der mit Essigsäure versetzt war, statt.
2.22 Eindeckeln
Die Hirnschnitte wurden auf gelatinisierten Objektträgern aufgezogen und
anschließend getrocknet. In einer aufsteigenden Alkoholreihe (70%, 96% und
100%) wurden die Präparate dehydriert, zweimal mit Xylol gespült, bevor mit
DEPEX („Serva, Feinbiochemie GmbH & Co“, Deutschland) eingedeckelt wurde.

Material und Methoden
41
2.23 Auswertung
Die Auswertung erfolgte an einem Mikroskop der Firma Olympus, Typ BH 2,
welches mit einem Bildanalysesystem („analysis“, Deutschland) verbunden war.
2.23.1 Qualitative Auswertungen
Bei den qualitativen Auswertungen wurden die Strukturen untersucht, welche die
einzelnen pAK im tectofugalen System markierten. Festgestellt wurden dabei die
Lokalisationen der Färbungen, als auch qualitative Unterschiede somatischer-
oder Neuropil-Markierungen.
2.23.2 Quantitative Auswertungen
Die quantitative Auswertung erfolgte bei neun Tauben in zwei Kerngebieten des
tectofugalen Systems, dem Nucleus subpraetectalis (SP) und dem Nucleus
interstitio-praetecto-subpraetectalis (IPS). Die Datenerhebung fand in der
gesamten rostro-caudal Ausdehnung des SP/IPS-Komplexes statt (A4.25–A.5.25,
Karten und Hodos 1967).
Von den Antikörpern gegen GAD, GABAA-Rezeptor, GluR4 und Parvalbumin
wurden die relative Zellzahl immunpositiver Zellen im SP und IPS bestimmt. Dazu
wurden im SP und IPS zwei Felder gleicher Größe (280µm x 220µm) angelegt. Ein
Feld war ventral, das andere im dorsalen Bereich der Kerne lokalisiert. Um den
prozentualen Anteil der markierten Neuronen zu bestimmen, wurden die gleichen
Felder in den Kresylviolettschnitten derselben Taube angelegt.
Eine statistische Analyse aller quantitativen Daten erfolgte mittels der Sofware
„Statistica for windows“ (Version 4.5, StatSoft, 1993, USA). Mögliche quantitative
Unterschiede in der Verteilung der immunpositiven Neuronen in der caudo-
rostralen Ausdehnung des Kernes sowie zwischen beiden Hemisphären wurden
mittels einer „Zwei-Wege ANOVA“ mit anschließender „post-hoc Untersuchung
(Scheffé-Test)“ durchgeführt.

Ergebnisse
42
3 Ergebnisse
Gegenstand der vorliegenden Untersuchungen war ein zum visuellen System
gehörendes Kerngebiet, der Nucleus subpraetectalis (SP), in der Taube Columba
livia. Mit Hilfe von Tracing-Experimenten mit den beiden Tracern Biotin-Dextran-
Amin (BDA) und der ? -Untereinheit des Choleratoxins (CtB) wurden die
Konnektivitäten des SP zu weiteren Hirnstrukturen festgestellt. Desweiteren
erfolgte eine immunhistochemische Charakterisierung der Expression von
Glutamatdecarboxylase, das den Neurotransmitter ? -Aminobuttersäure (GABA)
synthetisierende Enzym, von GABA- und Glutamat-Rezeptoren sowie des
Calcium-bindenden Proteins Parvalbumin im Nucleus subpraetectalis. Hierdurch
sollte der Einfluß des Nucleus subpraetectalis auf die durch die Tracing Daten
erkannten neuronalen verbundenen Strukturen näher beschrieben werden.
3.1 Tracing
Beim Tracing werden molekulare Substanzen, Tracer genannt, gezielt in
Hirnregionen injiziiert. Die Tracer werden durch Endocytose von den Neuronen
aufgenommen und entlang der Fortsätze dieser Zellen, den Axonen und
Dendriten, aktiv und passiv transportiert. Die molekularen Eigenschaften der
Tracer bestimmen hierbei die Laufeigenschaften, daß heißt die Geschwindigkeit
und die vorwiegende Richtung (anterograd – retrograd) des Transportes. Ebenfalls
abhängig vom Tracer erfolgt die anschließende Detektion der Signale, die nach
mikroskopischer Auswertung Aufschluß über die Konnektivitäten der einzelnen
Kerne gibt. Bei dieser Arbeit wurden die Tracer BDA und CtB in den
mesencephalen Nucleus subpraetectalis (SP) appliziert, wobei genaue
Injektionsorte für jede Taube einzeln mittels des histologischen Taubenatlas nach
Karten und Hodos (1967) bestimmt wurden (Tabelle 3).
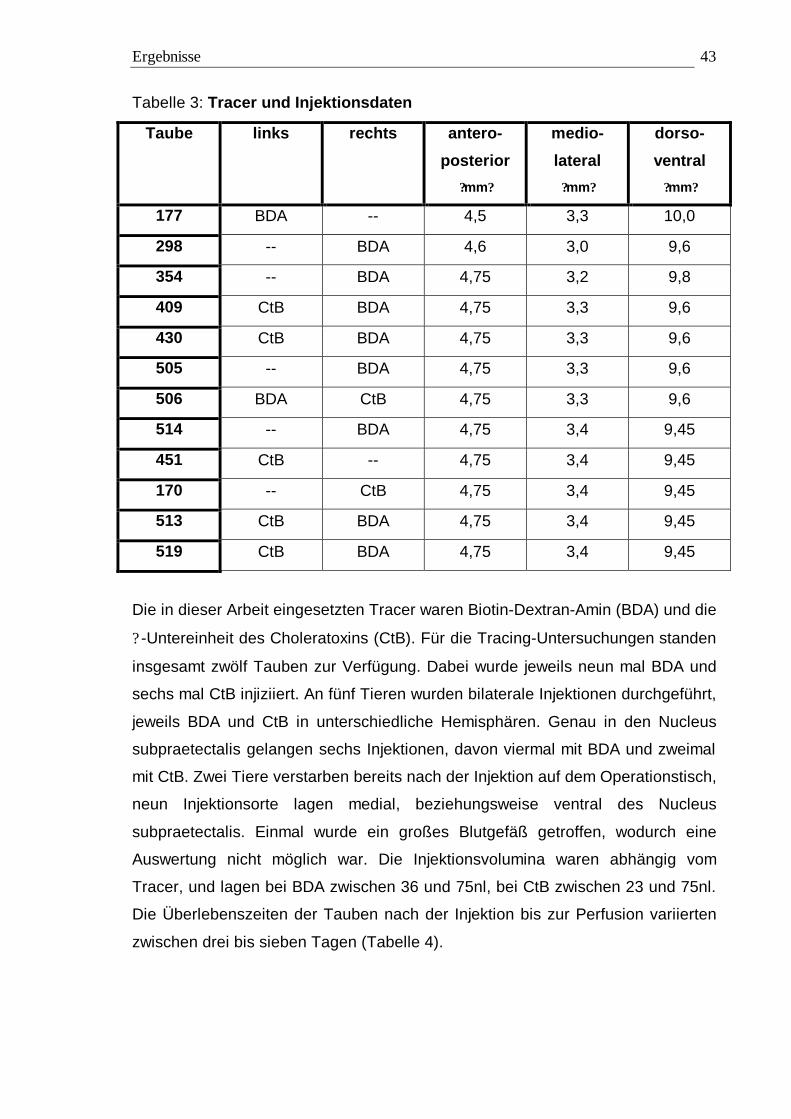
Ergebnisse
43
Tabelle 3: Tracer und Injektionsdaten
Taube links rechts antero-
posterior
?mm?
medio-
lateral
?mm?
dorso-
ventral
?mm?
177 BDA -- 4,5 3,3 10,0
298 -- BDA 4,6 3,0 9,6
354 -- BDA 4,75 3,2 9,8
409 CtB BDA 4,75 3,3 9,6
430 CtB BDA 4,75 3,3 9,6
505 -- BDA 4,75 3,3 9,6
506 BDA CtB 4,75 3,3 9,6
514 -- BDA 4,75 3,4 9,45
451 CtB -- 4,75 3,4 9,45
170 -- CtB 4,75 3,4 9,45
513 CtB BDA 4,75 3,4 9,45
519 CtB BDA 4,75 3,4 9,45
Die in dieser Arbeit eingesetzten Tracer waren Biotin-Dextran-Amin (BDA) und die
? -Untereinheit des Choleratoxins (CtB). Für die Tracing-Untersuchungen standen
insgesamt zwölf Tauben zur Verfügung. Dabei wurde jeweils neun mal BDA und
sechs mal CtB injiziiert. An fünf Tieren wurden bilaterale Injektionen durchgeführt,
jeweils BDA und CtB in unterschiedliche Hemisphären. Genau in den Nucleus
subpraetectalis gelangen sechs Injektionen, davon viermal mit BDA und zweimal
mit CtB. Zwei Tiere verstarben bereits nach der Injektion auf dem Operationstisch,
neun Injektionsorte lagen medial, beziehungsweise ventral des Nucleus
subpraetectalis. Einmal wurde ein großes Blutgefäß getroffen, wodurch eine
Auswertung nicht möglich war. Die Injektionsvolumina waren abhängig vom
Tracer, und lagen bei BDA zwischen 36 und 75nl, bei CtB zwischen 23 und 75nl.
Die Überlebenszeiten der Tauben nach der Injektion bis zur Perfusion variierten
zwischen drei bis sieben Tagen (Tabelle 4).
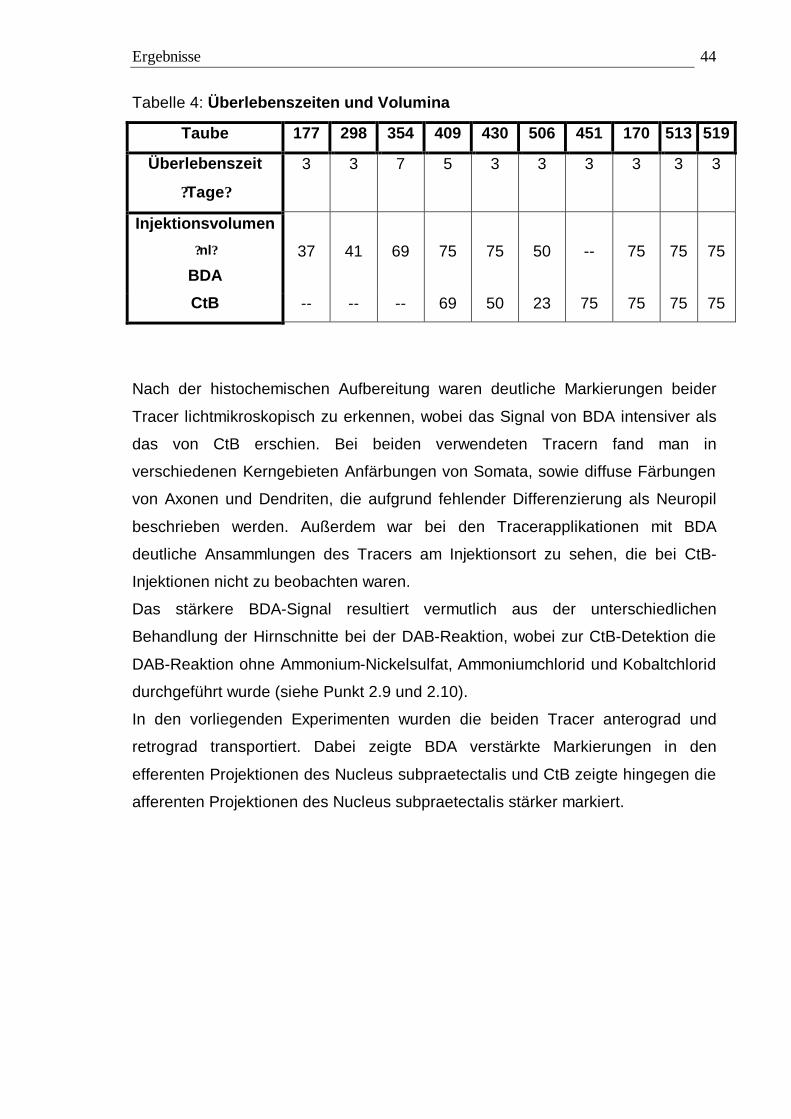
Ergebnisse
44
Tabelle 4: Überlebenszeiten und Volumina
Taube 177 298 354 409 430 506 451 170 513 519
Überlebenszeit
?Tage? 3 3 7 5 3 3 3 3 3 3
Injektionsvolumen
?nl?
BDA
37
41
69
75
75
50
--
75
75
75
CtB -- -- -- 69 50 23 75 75 75 75
Nach der histochemischen Aufbereitung waren deutliche Markierungen beider
Tracer lichtmikroskopisch zu erkennen, wobei das Signal von BDA intensiver als
das von CtB erschien. Bei beiden verwendeten Tracern fand man in
verschiedenen Kerngebieten Anfärbungen von Somata, sowie diffuse Färbungen
von Axonen und Dendriten, die aufgrund fehlender Differenzierung als Neuropil
beschrieben werden. Außerdem war bei den Tracerapplikationen mit BDA
deutliche Ansammlungen des Tracers am Injektionsort zu sehen, die bei CtB-
Injektionen nicht zu beobachten waren.
Das stärkere BDA-Signal resultiert vermutlich aus der unterschiedlichen
Behandlung der Hirnschnitte bei der DAB-Reaktion, wobei zur CtB-Detektion die
DAB-Reaktion ohne Ammonium-Nickelsulfat, Ammoniumchlorid und Kobaltchlorid
durchgeführt wurde (siehe Punkt 2.9 und 2.10).
In den vorliegenden Experimenten wurden die beiden Tracer anterograd und
retrograd transportiert. Dabei zeigte BDA verstärkte Markierungen in den
efferenten Projektionen des Nucleus subpraetectalis und CtB zeigte hingegen die
afferenten Projektionen des Nucleus subpraetectalis stärker markiert.

Ergebnisse
45
3.1.1 BDA
Es gelang in allen untersuchten Tieren eine gute Darstellung des Tracers BDA im
visuellen System der Taube. Hierbei war der Nachweis des Injektionsortes anhand
der zum Teil im Nucleus subpraetectalis verbleibenden Tracersubstanz möglich.
Injiziert wurde zwei mal in den ventro-lateralen Nucleus subpraetectalis, einmal in
den ventralen sowie einmal in den lateralen Nucleus subpraetectalis. Zusätzlich zu
den Tracerrückständen erkannte man im Nucleus subpraetectalis zumeist
vereinzelt auftretende Somata und Neuropilmarkierungen (Abbildung 1a,b). In
Abhängigkeit vom Injektionsort wurden die im folgenden beschriebenen
Konnektivitäten detektiert.
Nach SP-Injektionen zeigte der Nucleus interstitio-praetecto-subpraetectalis (IPS)
vor allem Fasermarkierung und zum Teil markierte Somata (Abbildung 1a,b). In
dem aus fünfzehn Schichten bestehenden Tectum opticum (TO) wurde vor allem
in der multipolaren Schicht 13 intensive Somata und Neuropil Markierung
beobachtet (Abbildung 2a,b). Der diencephale Nucleus rotundus (RT) zeigte eine
ausgeprägte Markierung von Fasern und Terminalien, somatische Markierungen
blieben aus, wobei sich von caudal nach rostral eine abnehmende dorso-mediale
Aussparung der Markierungen im Nucleus rotundus, bis zur vollständigen
Markierung des Kernes, zeigte (Abbildung 3a,b). Im Nucleus triangularis (T), der
dem Nucleus rotundus als dorsale Kappe aufliegt, fand man keine BDA-
Markierung von Somata oder Neuropil. Nach BDA-Injektionen in den Nucleus
subpraetectalis wurde der Nucleus praetectalis (PT) vor allem in den äußeren
Schichten markiert (Abbildung 4a,b). Es imponierten die Markierung von Fasern
und Terminalien, zum Teil waren auch somatische Markierungen zu erkennen.
Darüber hinaus wurden Fasern zwischen Nucleus subpraetectalis und Nucleus
rotundus, sowie zwischen Nucleus subpraetectalis und Nucleus praetectalis
markiert.
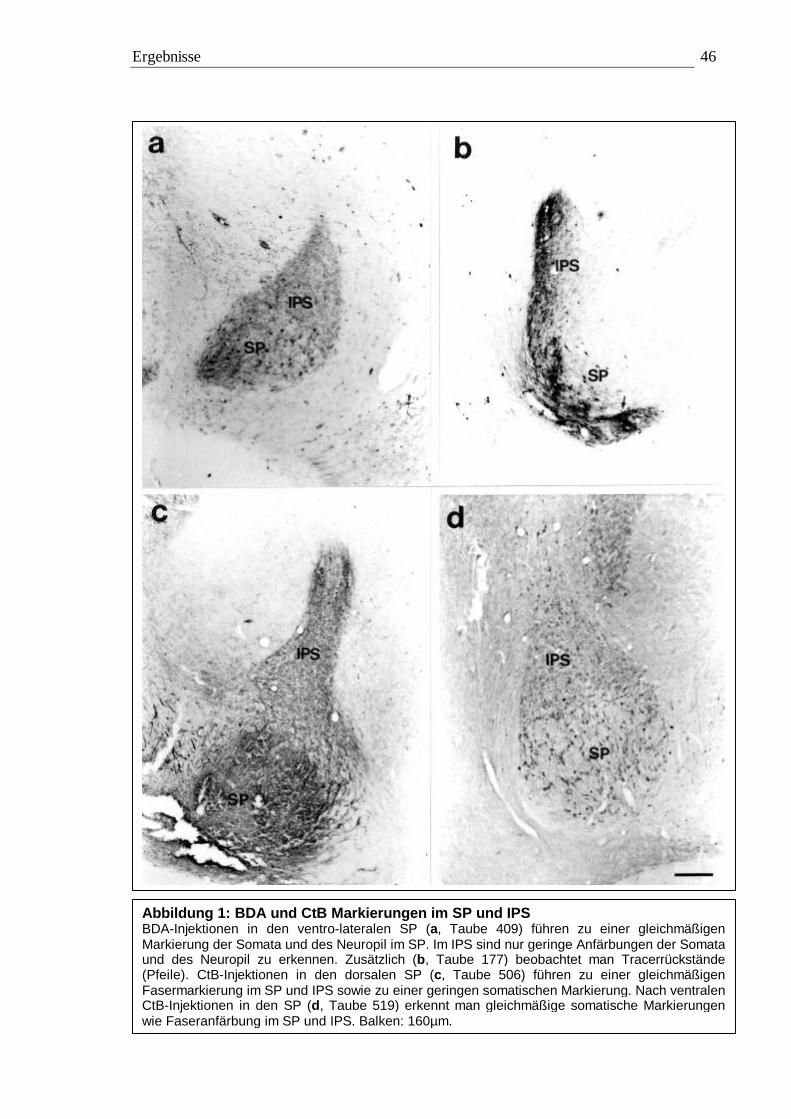
Ergebnisse
46
Abbildung 1: BDA und CtB Markierungen im SP und IPS BDA-Injektionen in den ventro-lateralen SP (a, Taube 409) führen zu einer gleichmäßigen Markierung der Somata und des Neuropil im SP. Im IPS sind nur geringe Anfärbungen der Somata und des Neuropil zu erkennen. Zusätzlich (b, Taube 177) beobachtet man Tracerrückstände (Pfeile). CtB-Injektionen in den dorsalen SP (c, Taube 506) führen zu einer gleichmäßigen Fasermarkierung im SP und IPS sowie zu einer geringen somatischen Markierung. Nach ventralen CtB-Injektionen in den SP (d, Taube 519) erkennt man gleichmäßige somatische Markierungen wie Faseranfärbung im SP und IPS. Balken: 160µm.
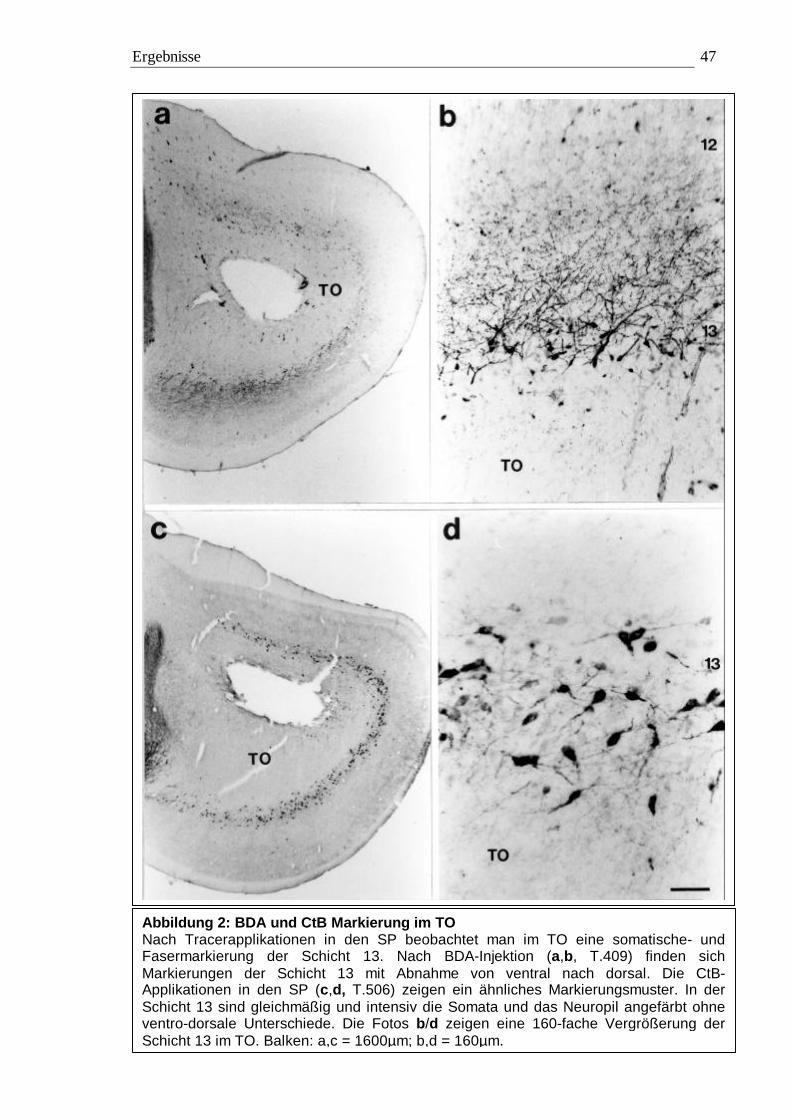
Ergebnisse
47
Abbildung 2: BDA und CtB Markierung im TO Nach Tracerapplikationen in den SP beobachtet man im TO eine somatische- und Fasermarkierung der Schicht 13. Nach BDA-Injektion (a,b, T.409) finden sich Markierungen der Schicht 13 mit Abnahme von ventral nach dorsal. Die CtB-Applikationen in den SP (c,d, T.506) zeigen ein ähnliches Markierungsmuster. In der Schicht 13 sind gleichmäßig und intensiv die Somata und das Neuropil angefärbt ohne ventro-dorsale Unterschiede. Die Fotos b/d zeigen eine 160-fache Vergrößerung der Schicht 13 im TO. Balken: a,c = 1600µm; b,d = 160µm.
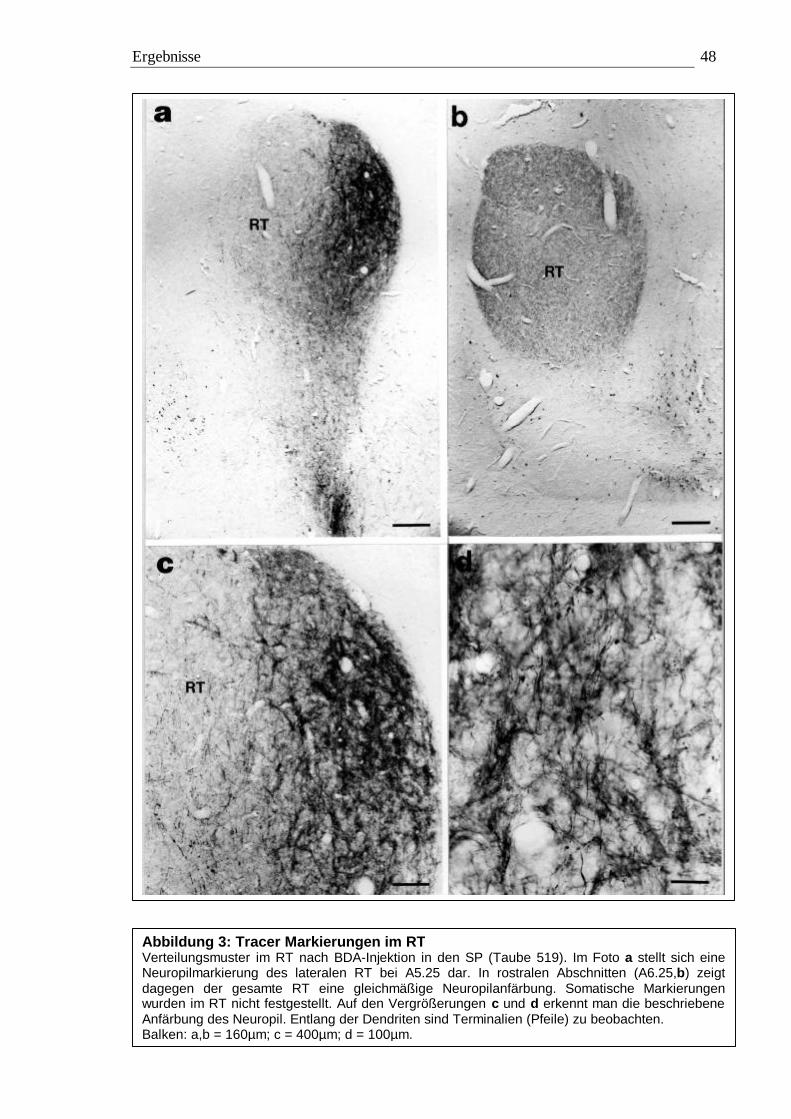
Ergebnisse
48
Abbildung 3: Tracer Markierungen im RT Verteilungsmuster im RT nach BDA-Injektion in den SP (Taube 519). Im Foto a stellt sich eine Neuropilmarkierung des lateralen RT bei A5.25 dar. In rostralen Abschnitten (A6.25,b) zeigt dagegen der gesamte RT eine gleichmäßige Neuropilanfärbung. Somatische Markierungen wurden im RT nicht festgestellt. Auf den Vergrößerungen c und d erkennt man die beschriebene Anfärbung des Neuropil. Entlang der Dendriten sind Terminalien (Pfeile) zu beobachten. Balken: a,b = 160µm; c = 400µm; d = 100µm.
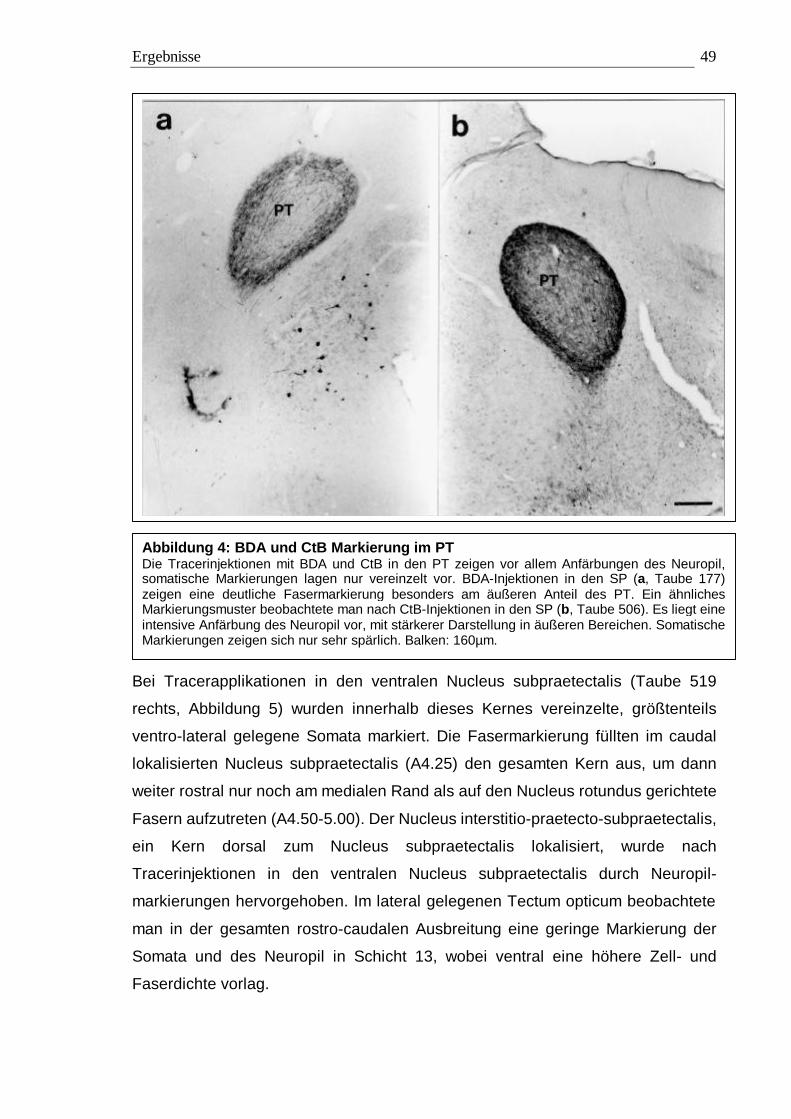
Ergebnisse
49
Bei Tracerapplikationen in den ventralen Nucleus subpraetectalis (Taube 519
rechts, Abbildung 5) wurden innerhalb dieses Kernes vereinzelte, größtenteils
ventro-lateral gelegene Somata markiert. Die Fasermarkierung füllten im caudal
lokalisierten Nucleus subpraetectalis (A4.25) den gesamten Kern aus, um dann
weiter rostral nur noch am medialen Rand als auf den Nucleus rotundus gerichtete
Fasern aufzutreten (A4.50-5.00). Der Nucleus interstitio-praetecto-subpraetectalis,
ein Kern dorsal zum Nucleus subpraetectalis lokalisiert, wurde nach
Tracerinjektionen in den ventralen Nucleus subpraetectalis durch Neuropil-
markierungen hervorgehoben. Im lateral gelegenen Tectum opticum beobachtete
man in der gesamten rostro-caudalen Ausbreitung eine geringe Markierung der
Somata und des Neuropil in Schicht 13, wobei ventral eine höhere Zell- und
Faserdichte vorlag.
Abbildung 4: BDA und CtB Markierung im PT Die Tracerinjektionen mit BDA und CtB in den PT zeigen vor allem Anfärbungen des Neuropil, somatische Markierungen lagen nur vereinzelt vor. BDA-Injektionen in den SP (a, Taube 177) zeigen eine deutliche Fasermarkierung besonders am äußeren Anteil des PT. Ein ähnliches Markierungsmuster beobachtete man nach CtB-Injektionen in den SP (b, Taube 506). Es liegt eine intensive Anfärbung des Neuropil vor, mit stärkerer Darstellung in äußeren Bereichen. Somatische Markierungen zeigen sich nur sehr spärlich. Balken: 160µm.
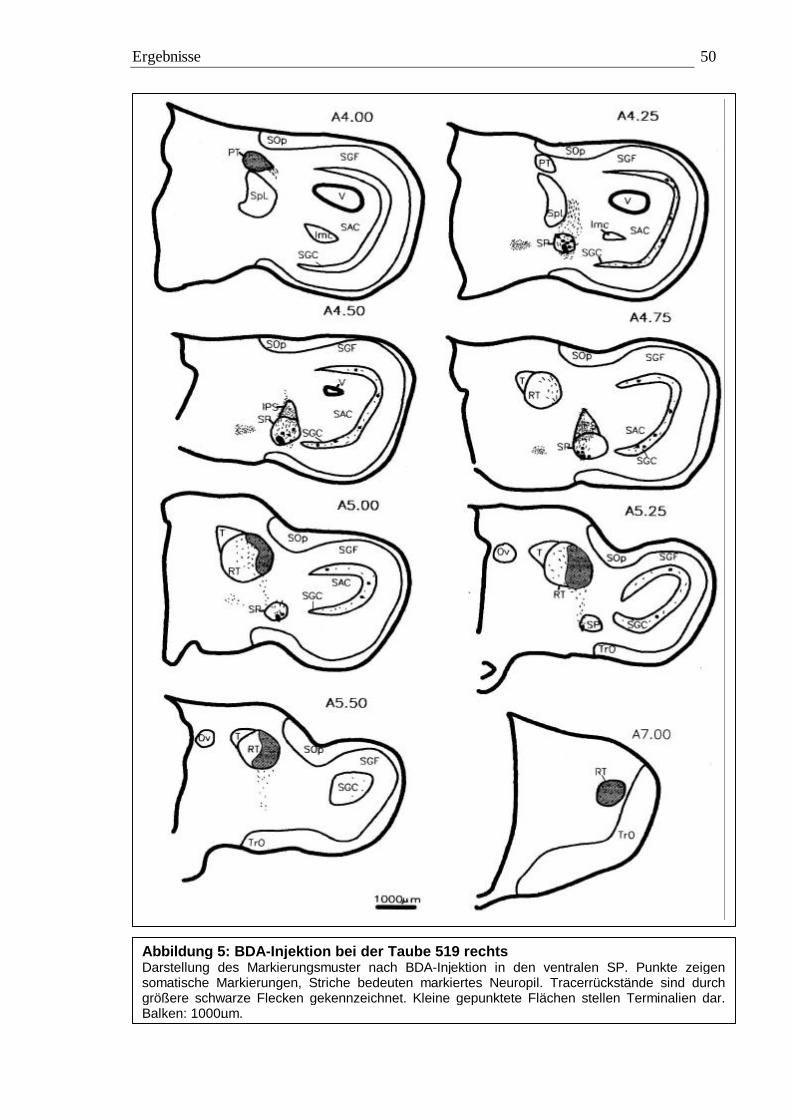
Ergebnisse
50
Abbildung 5: BDA-Injektion bei der Taube 519 rechts Darstellung des Markierungsmuster nach BDA-Injektion in den ventralen SP. Punkte zeigen somatische Markierungen, Striche bedeuten markiertes Neuropil. Tracerrückstände sind durch größere schwarze Flecken gekennzeichnet. Kleine gepunktete Flächen stellen Terminalien dar. Balken: 1000µm.

Ergebnisse
51
Der diencephale Nucleus rotundus stellte sich durch die ventrale BDA-Injektion in
den Nucleus subpraetectalis mit zahlreichen Terminalien und Fasern dar, es
wurden aber keine somatischen Markierungen beobachtet. Die Verteilung der
markierten Terminalien und Fasern im Nucleus rotundus betrafen caudal (A4.75)
nur den lateralen Nucleus wobei sich jedoch die Fasermarkierung weiter rostral
(A7.00) über den gesamten Kern erstreckte (Abbildung 3). Auch im Nucleus
praetectalis wurden Markierungen festgestellt, nachdem BDA in den ventralen
Nucleus subpraetectalis appliziert worden war. Der gesamte Nucleus praetectalis
war hierbei gekennzeichnet durch Markierungen von Terminalien, gefärbte Faser
wurden dabei, kreisförmig, in den äußeren Kernanteilen des Nucleus praetectalis
beobachtet.
Nach Injektion von BDA in den lateralen Nucleus subpraetectalis (Taube 513,
Abbildung 6), wurden im Nucleus subpraetectalis überwiegend
Neuropilmarkierung detektiert, welche im caudalen Bereich des Nucleus
subpraetectalis (A4.25-A4.75) auf den lateralen bis medialen Nucleus
subpraetectalis begrenzt waren, sich weiter rostral dagegen (A5.00-A5.25) auf den
gesamten Nucleus subpraetectalis ausdehnten. Somata wurden nur vereinzelt in
der Umgebung des Injektionsortes markiert. Der Nucleus interstitio-praetecto-
subpraetectalis imponierte durch zahlreiche Fasermarkierung, wobei auch hier im
caudalen (A4.50-4.75) Nucleus interstitio-praetecto-subpraetectalis die laterale
Hälfte des Nucleus interstitio-praetecto-subpraetectalis gefärbt war, und die
mediale Hälfte keine Markierung zeigte. Weiter rostral (A5.00) war dagegen der
gesamte Nucleus interstitio-praetecto-subpraetectalis markiert. Bei der lateralen
BDA-Injektion in den Nucleus subpraetectalis wurden im Tectum opticum Fasern
und zelluläre Strukturen in Schicht 13 markiert, mit einer vermehrten Markierung
im ventralen Abschnitt des Tectum opticum. Der Nucleus rotundus zeigte lateral
lokalisierte Fasern und Terminalien (A4.75-5.25), ohne BDA-Signal im Nucleus
triangularis. Die Ausdehnung der Markierung nahm nach rostral zu, bis der
gesamte Kern von Fasermarkierungen erfaßt wurde (A6.25). Somata wurden im
Nucleus rotundus nach BDA-Injektion in den Nucleus subpraetectalis nicht gefärbt.
Desweiteren wurden Fasern zwischen Nucleus subpraetectalis und Nucleus
rotundus dargestellt.
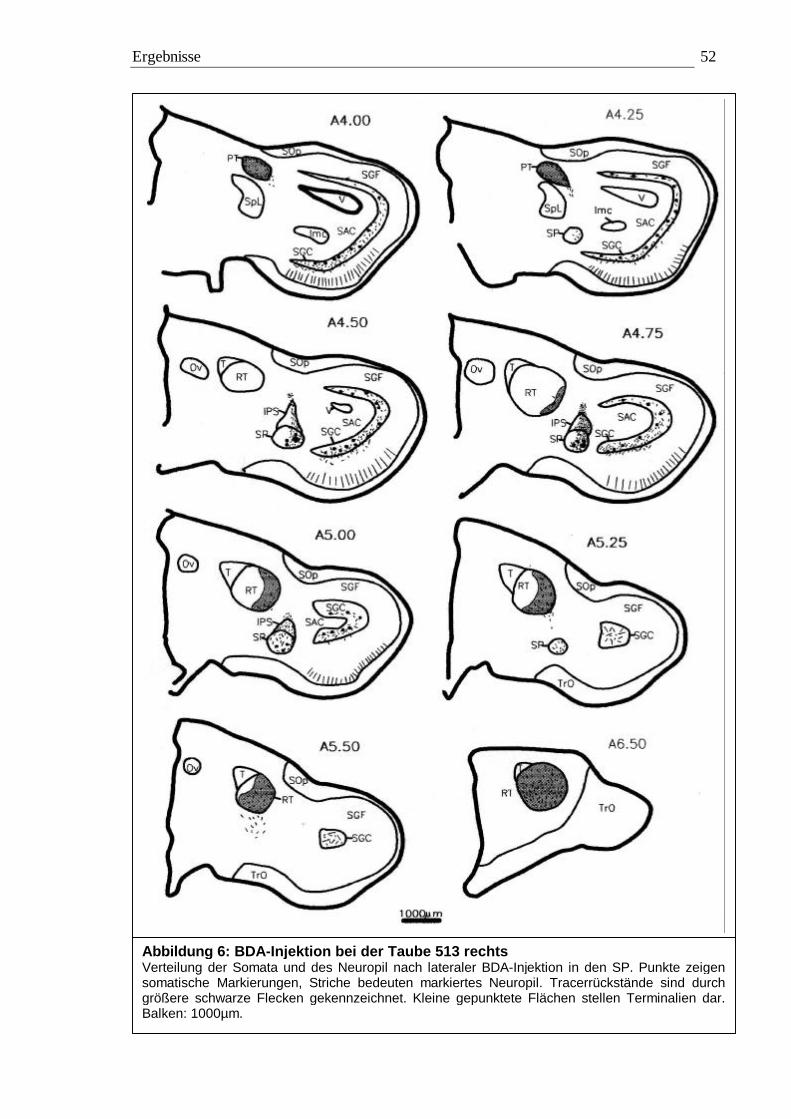
Ergebnisse
52
Abbildung 6: BDA-Injektion bei der Taube 513 rechts Verteilung der Somata und des Neuropil nach lateraler BDA-Injektion in den SP. Punkte zeigen somatische Markierungen, Striche bedeuten markiertes Neuropil. Tracerrückstände sind durch größere schwarze Flecken gekennzeichnet. Kleine gepunktete Flächen stellen Terminalien dar. Balken: 1000µm.

Ergebnisse
53
Nach der lateralen SP-Injektion wurden im gesamten Nucleus praetectalis
Terminalien markiert, wobei Fasermarkierungen auf den äußeren Nucleus
praetectalis begrenzt waren.
Bei zwei weiteren Tauben (Taube: 409 und 177, Abbildung 7 und 8) wurde der
Tracer BDA in den ventro-lateralen Abschnitt des Nucleus subpraetectalis injiziert.
Im caudalen (A4.25) Nucleus subpraetectalis waren Tracerrückstände zu sehen
(Abbildung 1a,b), weiter rostral hingegen wurden vermehrte Fasermarkierungen
und einzelne somatische Färbungen erkannt, begleitet von zahlreichen markierten
Terminalien. Die Verteilung der Markierungen von Somata, Neuropil und
Terminalien variierte innerhalb des Nucleus subpraetectalis. Im caudalen Nucleus
subpraetectalis waren vor allem Markierungen im ventro-lateralen Abschnitt zu
erkennen. Die Ausbreitung des BDA-Signals nach rostral nahm zu, bis in der
anterior-posterioren Ebene A5.00 gleichmäßig Somata, Neuropil und Terminalien
im Nucleus subpraetectalis markiert wurden. Der Nucleus interstitio-praetecto-
subpraetectalis zeigte schwache Markierungen von Somata und Fasern
(Abbildung 1a,b). In Schicht 13 des Tectum opticum wurde nach ventro lateraler
Injektion eine sehr intensive Somata- und Neuropilmarkierung beobachtet, wobei
im ventralen Tectum opticum die Dichte der Markierung höher war als im dorsalen
Tectum opticum. (Abbildung 2a,b). Auch bei den Tauben 409 und 177 waren
Fasern und Terminalien im Nucleus rotundus BDA-positiv, Somata aber nicht
markiert. Die Verteilung der BDA-Markierung im Nucleus rotundus zeigte von
caudal nach rostral auch bei diesen Tieren wieder eine zunächst laterale im
weiteren Verlauf zunehmende Markierung des Nucleus rotundus. Im Nucleus
triangularis wurde kein BDA-Signal erkannt. Zwischen Nucleus subpraetectalis
und Nucleus rotundus wurden wiederum BDA-positive Fasern detektiert. Der
Nucleus praetectalis imponierte durch Anfärbung der Terminalien und des
Neuropil mit dem Tracer BDA. Die Markierungen der Terminalien waren zumeist
im gesamten Nucleus praetectalis verteilt, mit Aussparungen im inneren (T.177,
A4.25), oder äußeren Anteil des Nucleus praetectalis (T.409, A4.50) (Abbildung
4a,b). Fasermarkierungen lagen vereinzelt vor, gehäuft am äußeren Rand des
Nucleus praetectalis. Auch in den Tauben 409 und 177 war das Neuropil im
Bereich zwischen Nucleus subpraetectalis und Nucleus praetectalis angefärbt.
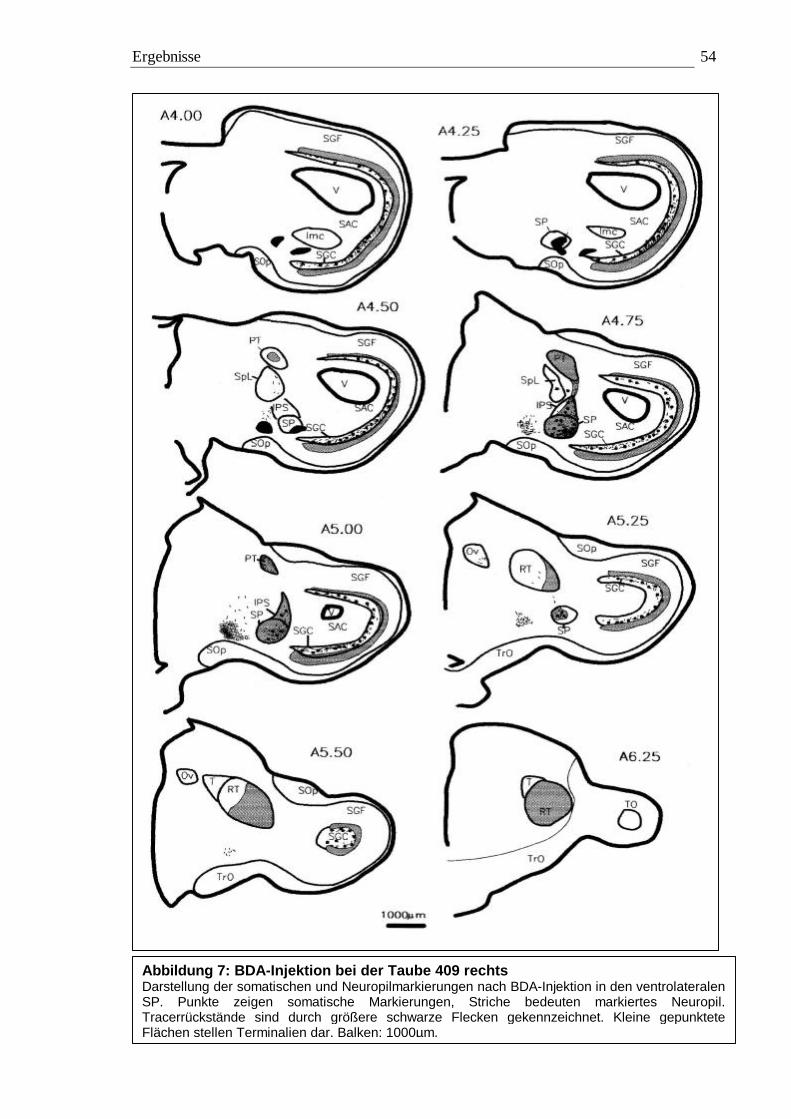
Ergebnisse
54
Abbildung 7: BDA-Injektion bei der Taube 409 rechts Darstellung der somatischen und Neuropilmarkierungen nach BDA-Injektion in den ventrolateralen SP. Punkte zeigen somatische Markierungen, Striche bedeuten markiertes Neuropil. Tracerrückstände sind durch größere schwarze Flecken gekennzeichnet. Kleine gepunktete Flächen stellen Terminalien dar. Balken: 1000µm.
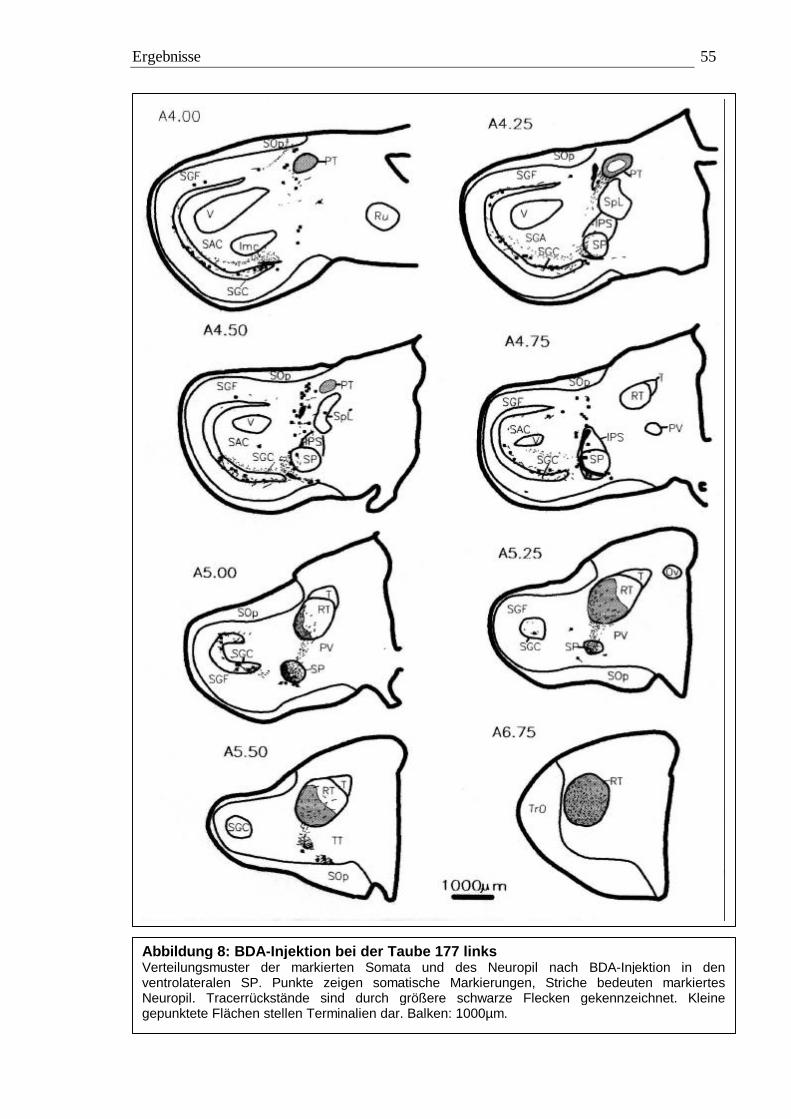
Ergebnisse
55
Abbildung 8: BDA-Injektion bei der Taube 177 links Verteilungsmuster der markierten Somata und des Neuropil nach BDA-Injektion in den ventrolateralen SP. Punkte zeigen somatische Markierungen, Striche bedeuten markiertes Neuropil. Tracerrückstände sind durch größere schwarze Flecken gekennzeichnet. Kleine gepunktete Flächen stellen Terminalien dar. Balken: 1000µm.

Ergebnisse
56
3.1.2 CtB
Eine erfolgreiche Darstellung des Tracers CtB nach den Injektionen in den
Nucleus subpraetectalis gelang bei zwei Tieren (Taube: 506 und 519), wobei in
der Taube 506 in den rechts hemisphärischen Nucleus subpraetectalis, in der
Taube 519 in den links hemisphärischen Nucleus subpraetectalis injiziert wurde.
Aufgrund der hohen Diffusionskapazität des Tracers CtB war der gesamte Kern
markiert. Eine weitere Subdifferenzierung des Injektionsortes war daher nicht
möglich.
Der Nucleus subpraetectalis zeigte bei beiden Tieren nach der CtB-Injektion eine
intensive Anfärbung des Neuropil und schwache somatische Markierungen
(Abbildung 1c,d). Im Nucleus interstitio-praetecto-subpraetectalis fand man jeweils
nach der Tracerapplikation in den Nucleus subpraetectalis eine starke Markierung
von Fasern, Somata wurden hingegen nur vereinzelt markiert (Abbildung 1c,d).
Das mesencephale Tectum opticum imponierte durch prominente Markierungen
der multipolaren Neuronen nebst Fortsätzen in der Schicht 13, wobei keine dorso-
ventralen Dichteunterschiede in der Markierung zu erkennen waren (Abbildung
2c,d).
Nach CtB-Injektion in den Nucleus subpraetectalis war im Nucleus rotundus das
Neuropil intensiv markiert, der Nucleus triangularis hingegen von Markierungen
ausgespart. Im dorsalen Nucleus praetectalis wurden CtB-positive Fasern und
vereinzelte CtB-positive Somata detektiert (Abbildung 4c). Zwischen dem Nucleus
subpraetectalis und dem Nucleus praetectalis wurden CtB-markierte Fasern mit
vereinzelt auftretenden CtB-positiven Somata erkannt.
Im Vergleich der einzelnen Tiere wurden Unterschiede nach den CtB-Injektionen
in den Nucleus subpraetectalis deutlich. Bei Taube 506 (Abbildung 9), wurde
vornehmlich das Neuropil und nur wenige Somata im Nucleus subpraetectalis
nach der Injektion erkannt. Die Ausdehnung der Fasermarkierungen variierte in
der rostro-caudal Achse, wobei nur der caudale Nucleus subpraetectalis (A4.25)
gleichmäßig und vollständig mit Fasern markiert war (Abbildung 1c). In der
medialen Ausdehnung des Nucleus subpraetectalis (A4.50-4.75) war die
Fasermarkierung hingegen auf den dorsalen Nucleus subpraetectalis begrenzt.
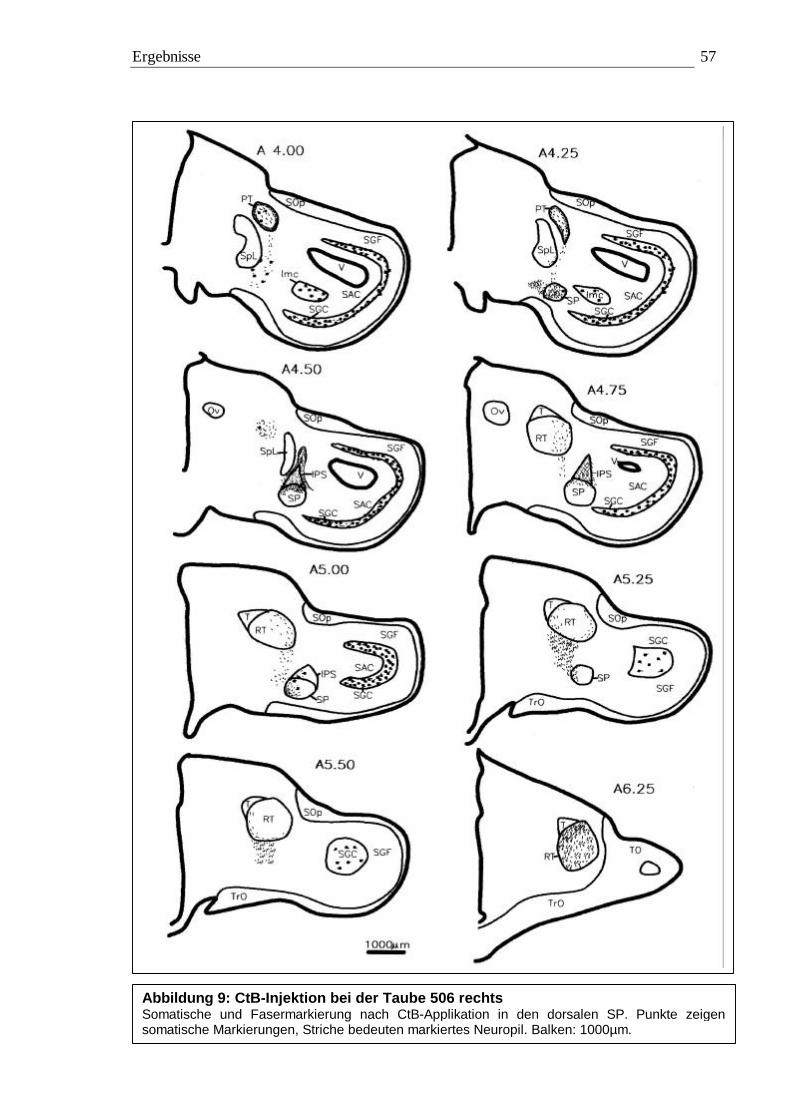
Ergebnisse
57
Abbildung 9: CtB-Injektion bei der Taube 506 rechts Somatische und Fasermarkierung nach CtB-Applikation in den dorsalen SP. Punkte zeigen somatische Markierungen, Striche bedeuten markiertes Neuropil. Balken: 1000µm.

Ergebnisse
58
Im rostralen Bereich des Kernes (A5.00-5.25) war das Neuropil zumeist im
medialen Anteil CtB-markiert, wobei die Faserrichtung auf den Nucleus rotundus
gerichtet war. Sehr vereinzelt waren in den rostralen Bereichen (A5.00) auch
Somata CtB-markiert.
Im Nucleus interstitio-praetecto-subpraetectalis waren zahlreiche CtB-positive
Fasern zu erkennen, die sich im caudalen Nucleus interstitio-praetecto-
subpraetectalis (A4.50-4.75) gleichmäßig verteilten (Abbildung 1c). Im rostralen
Abschnitt (A5.50) wurden moderat CtB-positive Somata und Neuropil deutlich.
Nach der Tracerapplikation in den Nucleus subpraetectalis zeigten sich im Tectum
opticum das Neuropil und die Somata in der Schicht 13 CtB-positiv (Abbildung
2c,d). Die CtB-Markierungen waren gleichmäßig in der dorso-ventralen
Ausdehnung dieser Schicht, ohne das Dichteunterschiede CtB-positiver Strukturen
erkannt wurden. Nur in der Taube 506 war eine leichte Abnahme der angefärbten
Somata und Fasern der Schicht 13 im rostralen Tectum opticum (A5.25-5.50) zu
beobachten.
Der diencephalen Nucleus rotundus imponierte durch CtB-positive Fasern, wo
hingegen keine Somata gefärbt waren. Das markierte Neuropil verteilte sich
primär (A4.75-5.00) im lateralen Nucleus rotundus, weiter rostral (bis A5.50) nahm
die laterale Markierung der Fasern ab. Gleichzeitig wurde eine Zunahme der CtB-
markierten Fasern im ventralen und medialen Nucleus rotundus erkannt. Noch
weiter rostral (A6.25) war dann der gesamte Kern mit Fasern und Terminalien
markiert.
Bei der Taube 506 zeigten sich im Nucleus praetectalis nach der rechts
hemisphärischen Injektion von CtB in den Nucleus subpraetectalis neben den
Markierungen von Fasern auch vereinzelte CtB-positive Somata, die auf den
zentralen Nucleus praetectalis begrenzt waren. Das Neuropil wurde hingegen im
gesamten Nucleus praetectalis CtB-markiert, wobei sich die Markierung in den
äußeren Bereichen des Kerns intensiver darstellte (Abbildung 4c). Auch bei der
Taube 506 wurden Fasermarkierungen zwischen Nucleus subpraetectalis und
Nucleus praetectalis erkannt.
Bei der Taube 519 (Abbildung 10) wurde CtB in den linken Nucleus
subpraetectalis appliziert. Im injizierten Kern wurden intensive, gleichmäßig
verteilte Fasermarkierungen und nur vereinzelt auftretende, CtB-positive Somata
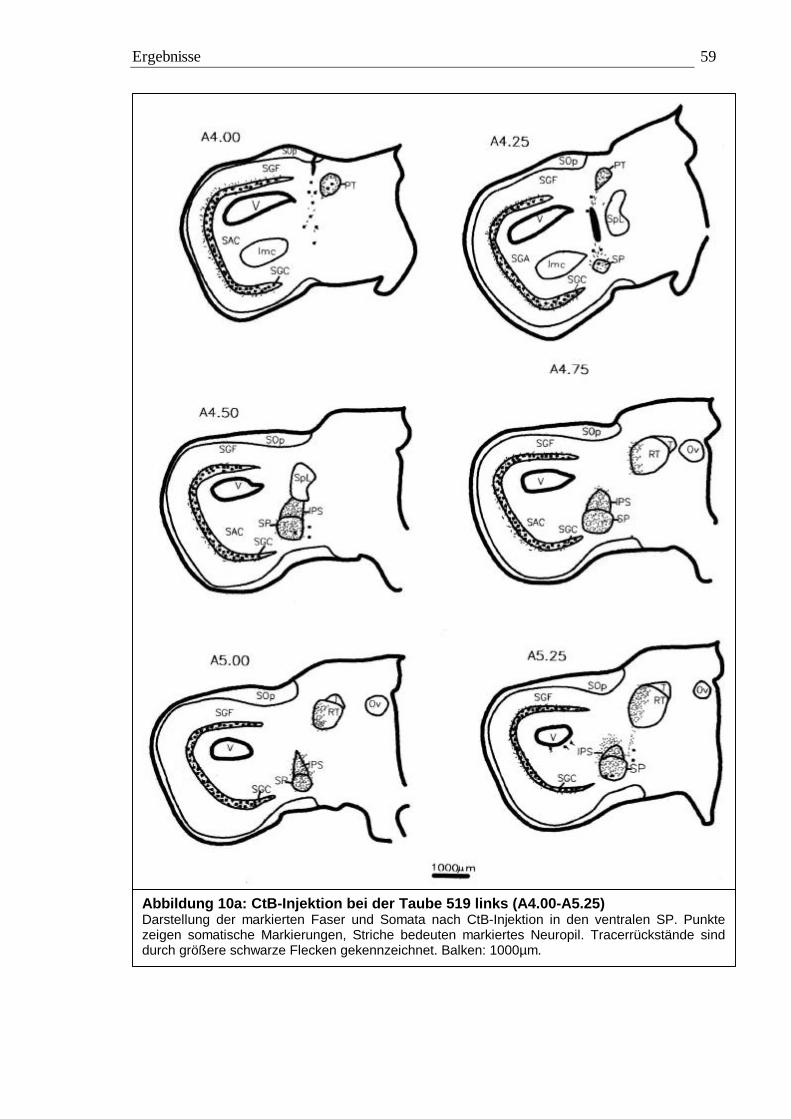
Ergebnisse
59
Abbildung 10a: CtB-Injektion bei der Taube 519 links (A4.00-A5.25) Darstellung der markierten Faser und Somata nach CtB-Injektion in den ventralen SP. Punkte zeigen somatische Markierungen, Striche bedeuten markiertes Neuropil. Tracerrückstände sind durch größere schwarze Flecken gekennzeichnet. Balken: 1000µm.
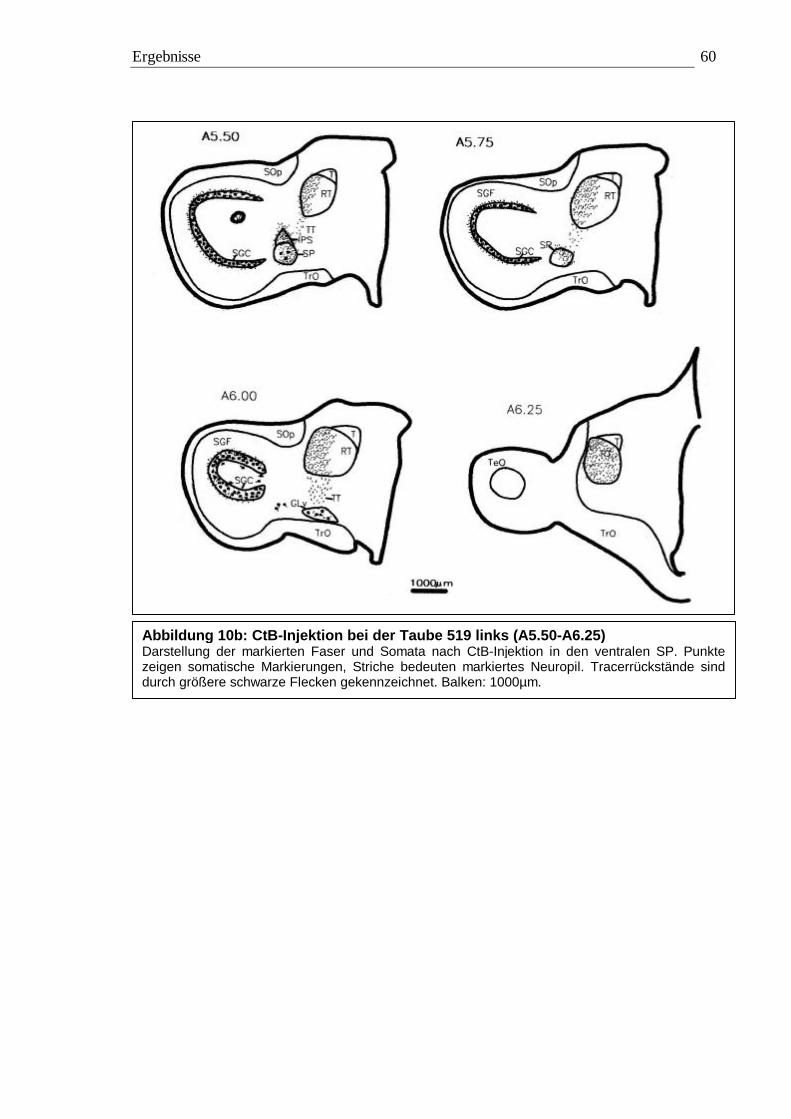
Ergebnisse
60
Abbildung 10b: CtB-Injektion bei der Taube 519 links (A5.50-A6.25) Darstellung der markierten Faser und Somata nach CtB-Injektion in den ventralen SP. Punkte zeigen somatische Markierungen, Striche bedeuten markiertes Neuropil. Tracerrückstände sind durch größere schwarze Flecken gekennzeichnet. Balken: 1000µm.

Ergebnisse
61
erkannt (Abbildung 1d), wobei im rostralen Nucleus subpraetectalis
Fasermarkierungen auf den medialen Anteil beschränkt blieben. CtB-positive
Fasern, nicht aber Somata, wurden im Nucleus interstitio-praetecto-
subpraetectalis mit einer gleichmäßigen Darstellung in der rostro-caudalen
Ausdehnung beobachtet (Abbildung 1d). Im Tectum opticum wurden intensive
Markierungen des Neuropil und der Somata, ohne dorso-ventrale
Dichteunterschiede, erkannt. Dieses Markierungsmuster entsprach dem von der
Taube 506. Zusätzlich zeigte die Schicht zehn CtB-markierte Fasern.
Bei der CtB-Injektion in den Nucleus subpraetectalis imponierte der Nucleus
rotundus durch eine homogene Fasermarkierung, ohne CtB-positive Somata,
wobei der Nucleus triangularis ausgespart blieb. Das markierte Neuropil
beschränkte sich primär auf den lateralen Nucleus rotundus, wobei von caudal
(A4.75) nach rostral (A6.25) eine verstärkte von lateral nach medial zunehmende
Fasermarkierung zu beobachten war von der schließlich der gesamte Kern
betroffen war. Zwischen Nucleus subpraetectalis und Nucleus rotundus waren
ebenfalls Fasern CtB-markiert.

Ergebnisse
62
Auch im Nucleus praetectalis wurde nach Tracerapplikation in den Nucleus
subpraetectalis CtB-positive Faser und Somata detektiert, wobei Somata zumeist
zentral im Nucleus praetectalis lagen, die Fasermarkierungen dagegen häufig den
gesamten Kern betrafen. Außerdem wurden zwischen Nucleus subpraetectalis
und Nucleus praetectalis vereinzelt Tracerrückstände, markierte Somata und
Neuropil erkannt. Die Tabellen 5 und 6 fassen die Markierungsmuster nach BDA-
und CtB-Injektionen und in den Nucleus subpraetectalis zusammen.
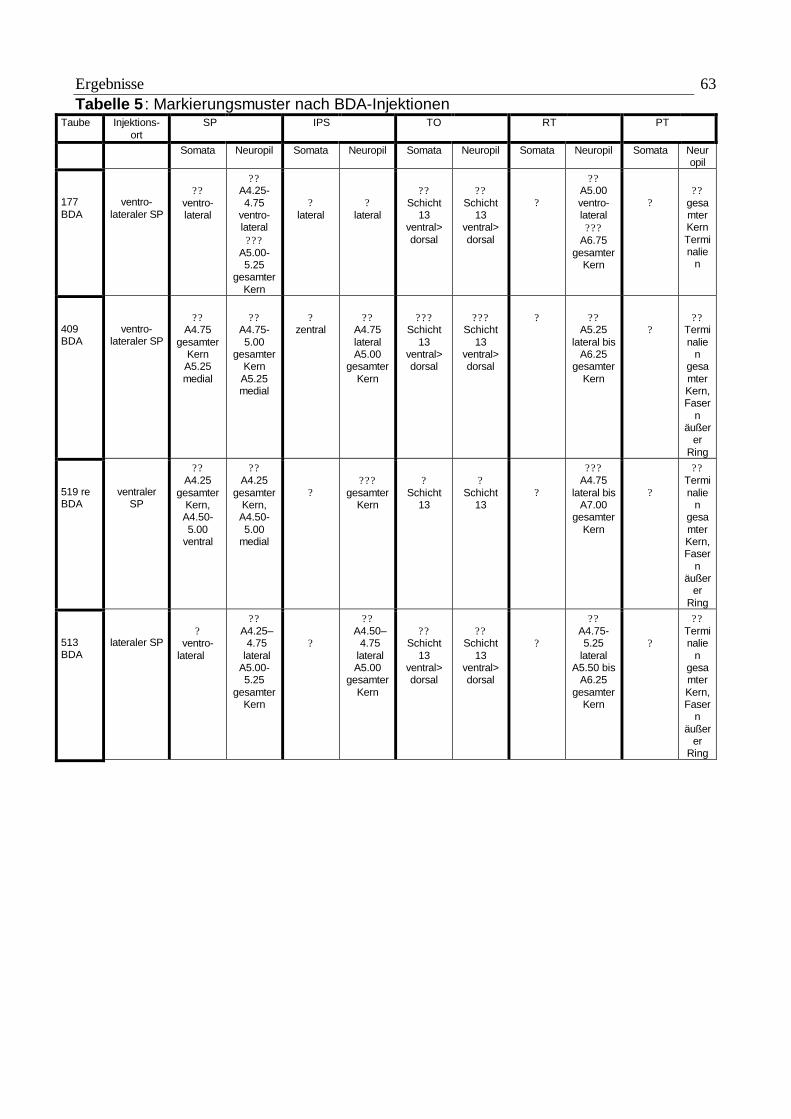
Ergebnisse
63
Tabelle 5 : Markierungsmuster nach BDA-Injektionen Taube Injektions-
ort SP IPS TO RT PT
Somata Neuropil Somata Neuropil Somata Neuropil Somata Neuropil Somata Neuropil
177 BDA
ventro-lateraler SP
? ?
ventro-lateral
? ? A4.25-4.75
ventro-lateral ? ? ?
A5.00-5.25
gesamter Kern
?
lateral
?
lateral
? ?
Schicht 13
ventral> dorsal
? ?
Schicht 13
ventral> dorsal
?
? ? A5.00 ventro-lateral ? ? ?
A6.75 gesamter
Kern
?
? ?
gesamter Kern Terminalie
n
409 BDA
ventro-lateraler SP
? ?
A4.75 gesamter
Kern A5.25 medial
? ?
A4.75-5.00
gesamter Kern A5.25 medial
?
zentral
? ?
A4.75 lateral A5.00
gesamter Kern
? ? ?
Schicht 13
ventral> dorsal
? ? ?
Schicht 13
ventral> dorsal
?
? ?
A5.25 lateral bis
A6.25 gesamter
Kern
?
? ?
Terminalie
n gesamter Kern, Faser
n äußer
er Ring
519 re BDA
ventraler SP
? ? A4.25
gesamter Kern, A4.50-5.00
ventral
? ? A4.25
gesamter Kern, A4.50-5.00
medial
?
? ? ?
gesamter Kern
?
Schicht 13
?
Schicht 13
?
? ? ? A4.75
lateral bis A7.00
gesamter Kern
?
? ? Terminalie
n gesamter Kern, Faser
n äußer
er Ring
513 BDA
lateraler SP
?
ventro-lateral
? ? A4.25–
4.75 lateral
A5.00-5.25
gesamter Kern
?
? ? A4.50–
4.75 lateral A5.00
gesamter Kern
? ?
Schicht 13
ventral> dorsal
? ?
Schicht 13
ventral> dorsal
?
? ? A4.75-5.25
lateral A5.50 bis
A6.25 gesamter
Kern
?
? ? Terminalie
n gesamter Kern, Faser
n äußer
er Ring

Ergebnisse
64
Tabelle 6: Markierungsmuster nach CtB-Injektionen Taube Injektions-
ort SP IPS TO RT PT
Somata Neuropil Somata Neuropil Somata Neuropil Somata Neuropil Somata Neuropil
519 li CtB
ventraler SP
? zentral
? ? A4.50-5.25 ventral A5.50 zentral
?
? ? ? gesamter IPS
? ? ? Schicht 13
? ? ? Schicht 13 ? ? Schicht 10
? ? ? A4.75-5.75 lateral A6.25 gesamter Kern
? A4.00 zentral
? ? A4.00 äußerer Ring A4.25 gesamter PT
506 CtB
dorsaler SP
? dorsaler SP
? ? A4.25 gesamter SP A4.50-4.75 dorsal A5.00-5.25 medial
? A5.00 medial
? ? A4.50-4.75 gesamter IPS A5.00 medial
? ? ? Schicht 13
? ? ? Schicht 13
? ? ? A4.75-5.00 Lateral A5.25 zentrale Aussparung A5.50 ventro-medial A6.25 gesamter RT
? A4.00 zentral
? ? gesamter PT besonders äußerer Ring
Legende ? keine Markierung ? kaum Markierung ? ? moderate Markierung ? ? ? starke Markierung

Ergebnisse
65
3.2 Immunhistochemie Die verwendeten polyklonalen und monoklonalen Antikörper waren gegen die
Antigene verschiedener Rezeptoruntereinheiten, Enzyme und Proteine gerichtet.
Dabei handelte es sich um die Glutamatrezeptoruntereinheit Glu R4, die ß-
Untereinheit des GABAA-Rezeptor, das den Neurotransmitter ? -Aminobuttersäure
synthetisierende Enzym Glutamatdecarboxylase (GAD) und das Calcium-
bindende Protein Parvalbumin. In der zunächst folgenden qualitativen Auswertung
werden die Immunmarkierungen dieser Antikörper in verschiedenen, dem
visuellen System der Taube zugehörigen Strukturen, wie dem Nucleus
subpraetectalis, dem Nucleus interstitio-praetecto-subpraetectalis, dem Tectum
opticum und dem Nucleus rotundus dargestellt. Bei der anschließenden
quantitativen Auswertung werden Unterschiede in der Häufigkeit der diese
Proteine exprimierenden Neuronen im Nucleus subpraetectalis und im Nucleus
interstitio-praetecto-subpraetectalis beschrieben.
3.2.1 Kontrollexperimente
Zur Überprüfung der Spezifität der sekundären Antikörper wurden
Kontrollexperimente durchgeführt. Bei diesen Kontrollen wurden die primären
Antikörper durch ein immunologisch nicht reaktives Serum ersetzt, während die
Immunhistochemie ansonsten unverändert blieb. Bei diesen Kontrollexperimenten
wurden weder somatische noch diffuse Markierungen beobachtet.
3.2.2 Qualitative Auswertung
Mit allen vier verwendeten Antikörpern wurde im Nucleus Subpraetectalis eine
gleichmäßig verteilte somatische Markierung erkannt (Abbildung 11a-d). Im
Gegensatz dazu führten nur die Antikörper gegen GAD, GABAA-Rezeptor und
Parvalbumin zu einer deutlichen Anfärbung des Neuropil, während mit den
Antikörpern gegen Glu R4 keine Neuropilmarkierung beobachtet wurde.
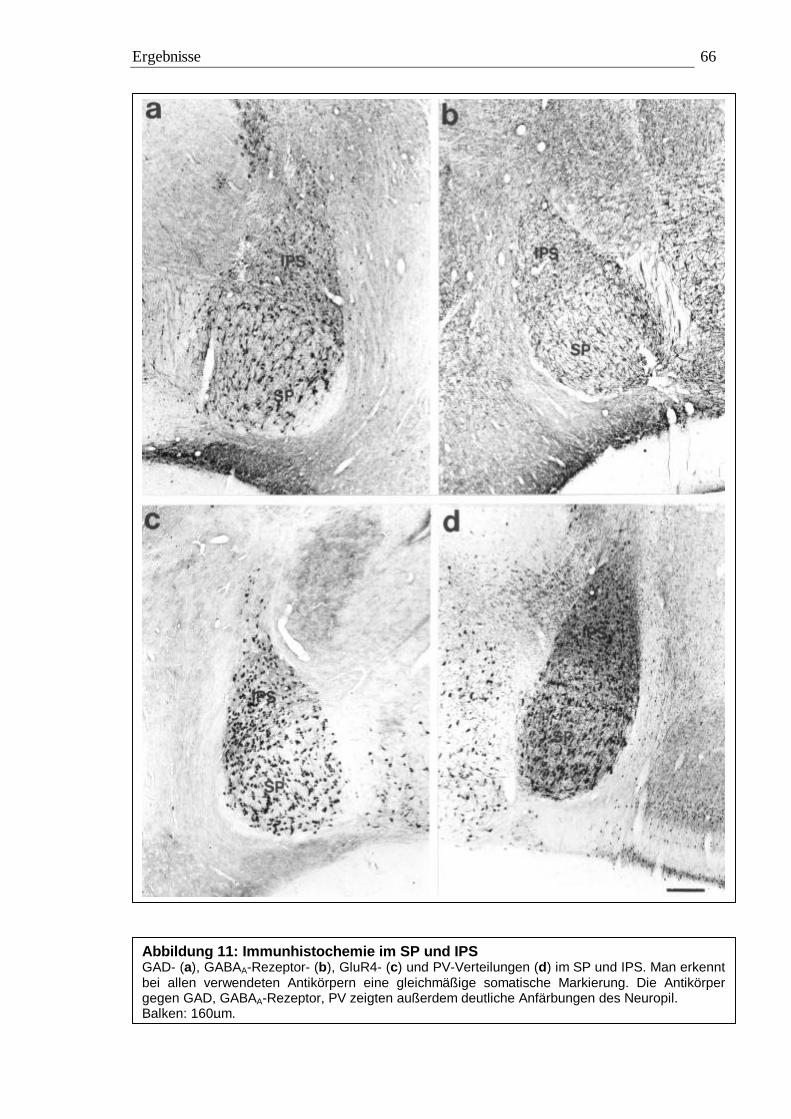
Ergebnisse
66
Abbildung 11: Immunhistochemie im SP und IPS GAD- (a), GABAA-Rezeptor- (b), GluR4- (c) und PV-Verteilungen (d) im SP und IPS. Man erkennt bei allen verwendeten Antikörpern eine gleichmäßige somatische Markierung. Die Antikörper gegen GAD, GABAA-Rezeptor, PV zeigten außerdem deutliche Anfärbungen des Neuropil. Balken: 160µm.

Ergebnisse
67
Im Nucleus interstitio-praetecto-subpraetectalis wurden intensive Antigen-
markierungen der verschiedenen Proteine beobachtet. Neben den somatischen
Markierungen aller verwendeten Antikörper in dieser mesencephalen Struktur war
auch das Neuropil im Nucleus interstitio-praetecto-subpraetectalis immunpositiv
gegen den GAD, GABAA-Rezeptor und Parvalbumin markiert (Abbildung 11a-d).
Die Immunhistochemie mit dem Glu R4-Antikörper zeigte dagegen ausschließlich
immunpositive Somata.
Im Tectum opticum wurden mit den eingesetzten primären Antikörpern gegen
GABAA-Rezeptor, GAD und Parvalbumin Somata und Neuropil markiert, wobei die
diversen detektierten Antigene Unterschiede in der Lokalisation und der Intensität
der Markierung zeigten (Abbildung 12a-d). Die GAD-Antikörper führten zu einer
starken Anfärbung der Somata in den Schichten 2,3 und 13, die Schichten 4, 5, 7
und 10 waren somatisch etwas schwächer markiert. Neuropilmarkierungen stellten
sich verstärkt in den superficialen Schichten 2, 3 und 4 dar (Abbildung 12a). Bei
Antikörpern gegen den GABAA-Rezeptor zeigten sich deutliche somatische
Markierungen in Schicht 13 sowie intensive Markierung des Neuropil in den
Schichten 2, 3, 4 und 5 sowie in der Schicht 13 (Abbildung 12b). In der
Immunhistochemie gegen Glu R4 zeigte sich keine somatische Anfärbung oder
Markierung des Neuropil im Tectum opticum (Abbildung 12c). Intensive
Parvalbumin-positive Somata wurden in den Schichten 2, 3, 4, 6 und 8 des
Tectum opticum beobachtet, etwas abgeschwächt lagen somatische
Markierungen in den Schichten 10 und 13 vor. Neuropilmarkierung beobachtete
man im Tectum opticum durch Parvalbumin-Antikörper deutlich in den Schichten
2, und 3, hingegen waren in den Schichten 4, 6 und 8 moderatere Markierungen
zu erkennen (Abbildung 12d).
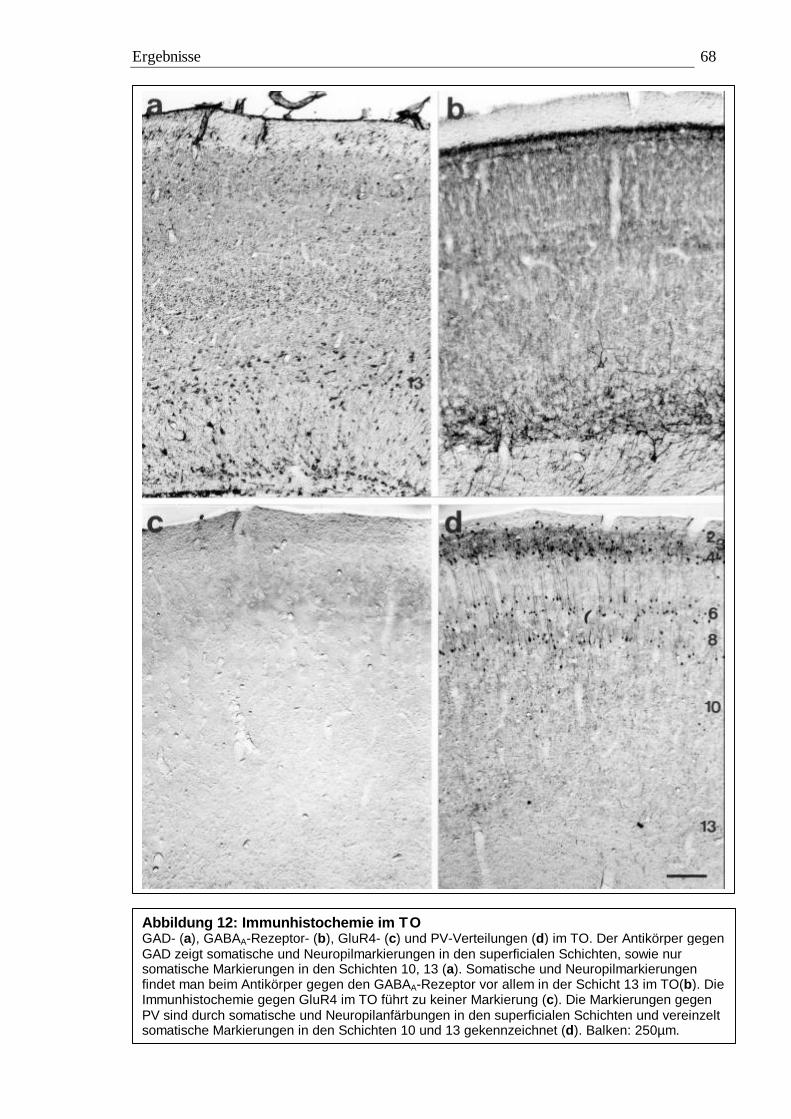
Ergebnisse
68
Abbildung 12: Immunhistochemie im TO GAD- (a), GABAA-Rezeptor- (b), GluR4- (c) und PV-Verteilungen (d) im TO. Der Antikörper gegen GAD zeigt somatische und Neuropilmarkierungen in den superficialen Schichten, sowie nur somatische Markierungen in den Schichten 10, 13 (a). Somatische und Neuropilmarkierungen findet man beim Antikörper gegen den GABAA-Rezeptor vor allem in der Schicht 13 im TO(b). Die Immunhistochemie gegen GluR4 im TO führt zu keiner Markierung (c). Die Markierungen gegen PV sind durch somatische und Neuropilanfärbungen in den superficialen Schichten und vereinzelt somatische Markierungen in den Schichten 10 und 13 gekennzeichnet (d). Balken: 250µm.

Ergebnisse
69
Im diencephalen Nucleus rotundus wurden mit allen primären Antikörpern
Neuropil-Markierungen erkannt, hingegen lagen somatische Markierungen nur
vereinzelt vor. Eine deutliche Darstellung des Neuropil wurde mit dem Antikörper
gegen GAD beobachtet bei nur schwacher Markierung der Somata (Abbildung
13a). Während der Antikörper gegen den GABAA-Rezeptor nur im Neuropil des
Nucleus rotundus erkannt wurde (Abbildung 13b) zeigten hingegen Antikörper
gegen die AMPA-Rezeptoruntereinheit Glu R4 eine deutliche Anfärbung der
Somata und des Neuropil (Abbildung 13c),. Das Calcium-bindenden Protein
Parvalbumin zeigte ebenso wie GAD eine schwache zelluläre Markierung, aber
eine intensive Färbung des Neuropil im Nucleus rotundus (Abbildung 13d).
Eine Übersicht über die qualitative Auswertung stellt Tabelle 7 dar.
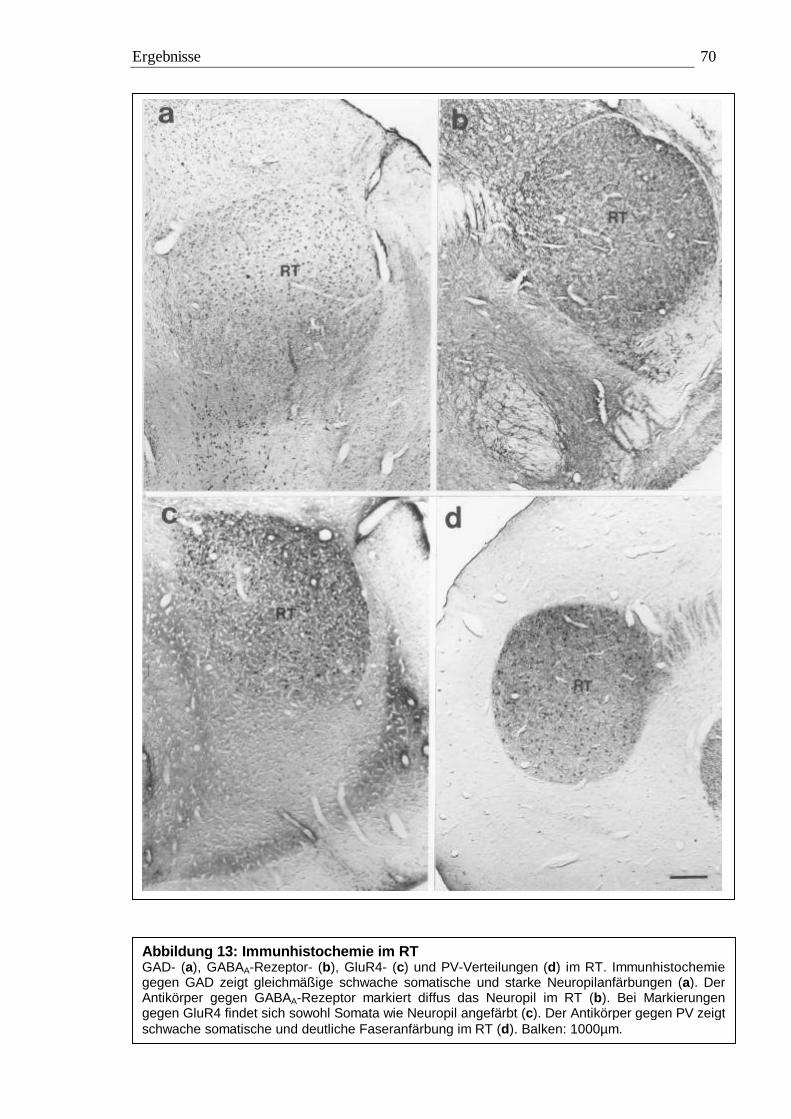
Ergebnisse
70
Abbildung 13: Immunhistochemie im RT GAD- (a), GABAA-Rezeptor- (b), GluR4- (c) und PV-Verteilungen (d) im RT. Immunhistochemiegegen GAD zeigt gleichmäßige schwache somatische und starke Neuropilanfärbungen (a). Der Antikörper gegen GABAA-Rezeptor markiert diffus das Neuropil im RT (b). Bei Markierungen gegen GluR4 findet sich sowohl Somata wie Neuropil angefärbt (c). Der Antikörper gegen PV zeigt schwache somatische und deutliche Faseranfärbung im RT (d). Balken: 1000µm.
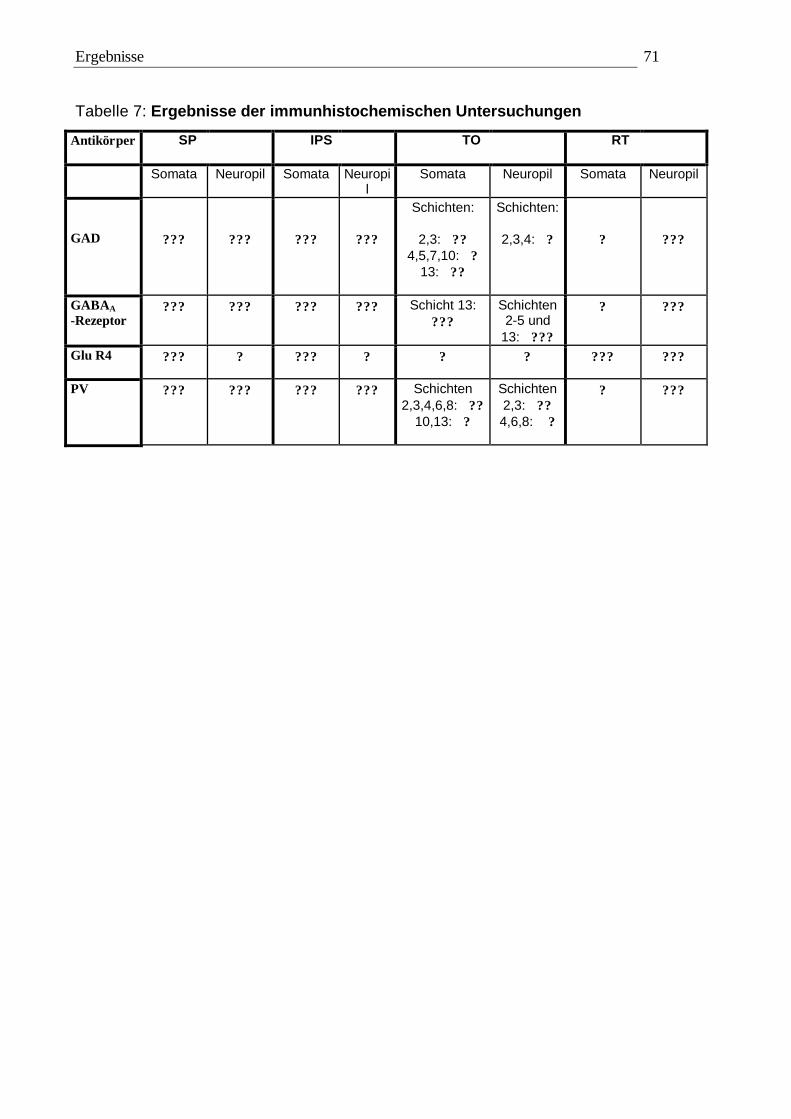
Ergebnisse
71
Tabelle 7: Ergebnisse der immunhistochemischen Untersuchungen
Antikörper
SP IPS TO RT
Somata Neuropil Somata Neuropil
Somata
Neuropil Somata Neuropil
GAD
? ? ?
? ? ?
? ? ?
? ? ?
Schichten:
2,3: ? ? 4,5,7,10: ?
13: ? ?
Schichten:
2,3,4: ?
?
? ? ?
GABAA -Rezeptor
? ? ? ? ? ? ? ? ? ? ? ? Schicht 13: ? ? ?
Schichten 2-5 und
13: ? ? ?
? ? ? ?
Glu R4
? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?
PV
? ? ? ? ? ? ? ? ? ? ? ? Schichten 2,3,4,6,8: ? ?
10,13: ?
Schichten 2,3: ? ? 4,6,8: ?
? ? ? ?

Ergebnisse
72
3.2.3 Quantitative Auswertung
Die quantitative Analyse immunpositiver Neuronen und Kresylviolett gefärbter
Neuronen innerhalb des Nucleus subpraetectalis und des Nucleus interstitio-
praetecto-subpraetectalis, ermöglichte die Bestimmung des prozentualen Anteils
GAD-, GABAA-Rezeptor-, Parvalbumin- und Glu R4-positiver Zellen in den beiden
Kerngebieten. Die quantitativen Analysen im Nucleus subpraetectalis erstreckte
sich über die caudo-rostrale Ausdehnung dieser Struktur von A4.25 bis A 5.25
(Anatomischer Atlas der Taube, Karten & Hodos, 1967). Die Zellzählung im
Nucleus interstitio-praetecto-subpraetectalis wurde in den caudo-rostralen
Bereichen zwischen A 4.50-A5.00 durchgeführt. Bei der Bestimmung des
prozentualen Anteils immunpositiver Neurone wurden die Daten aus beiden
tectalen Hälften gemittelt, da keine offensichtlichen Unterschiede zwischen linkem
und rechtem Nucleus subpraetectalis und Nucleus interstitio-praetecto-
subpraetectalis beobachtet wurden (siehe statistische Auswertung, 3.2.3.1 und
3.2.3.2). Die statistische Analyse wurde mittels der Sofware „Statistica for
windows“ (Version 4.5, StatSoft, 1993, USA) durchgeführt. Mittels einer „Zwei-
Wege ANOVA“ mit anschließender „post-hoc Untersuchung (Scheffé-Test)“
erfolgte die Bestimmung quantitativer Unterschiede in der Verteilung der
immunpositiven Neuronen in der caudo-rostralen Ausdehnung des Kernes sowie
zwischen beiden Hemisphären.
3.2.3.1 Quantitative Auswertung im Nucleus subpraetectalis
Die quantitativen Auswertungen immunpositiver Neuronen im Nucleus
subpraetectalis zeigten, daß eine Vielzahl der Nervenzellen dieses praetectalen
Kernes die untersuchten Antigene exprimierten. Bei den Antiköpern gegen GAD,
GABAA-Rezeptor und Parvalbumin imponierte insgesamt eine konstante
Verteilung der markierten Neurone in der caudo-rostralen Ausdehnung des
Nucleus subpraetectalis (Abbildungen 14 und 15). Der Antikörper gegen Glu R4
hingegen zeigte in der caudo-rostralen Ausdehnung ansteigende Werte
immunpositiver Nervenzellen, während links-rechts Unterschiede zwischen beiden
Hemisphären nicht deutlich wurden (Abbildungen 14 und 15).
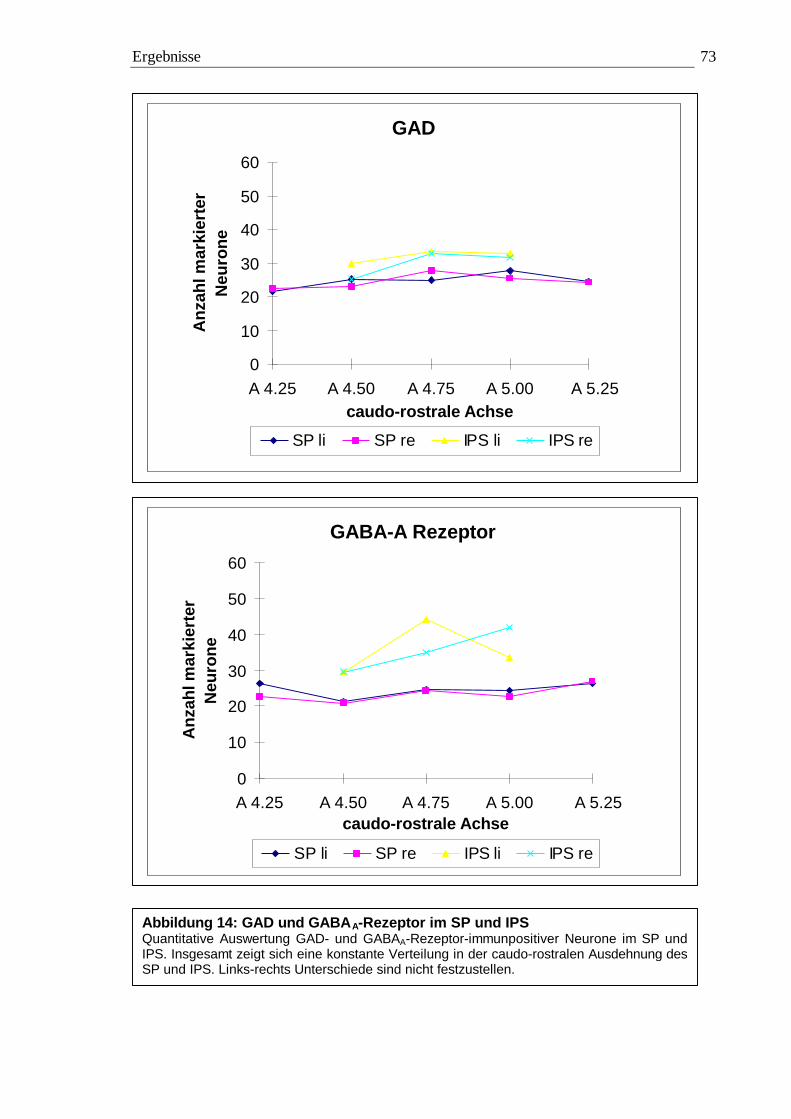
Ergebnisse
73
GAD
0
10
20
30
40
50
60
A 4.25 A 4.50 A 4.75 A 5.00 A 5.25caudo-rostrale Achse
Anz
ahl m
arki
erte
r N
euro
ne
SP li SP re IPS li IPS re
GABA-A Rezeptor
0
10
20
30
40
50
60
A 4.25 A 4.50 A 4.75 A 5.00 A 5.25caudo-rostrale Achse
Anz
ahl m
arki
erte
r N
euro
ne
SP li SP re IPS li IPS re
Abbildung 14: GAD und GABAA-Rezeptor im SP und IPS Quantitative Auswertung GAD- und GABAA-Rezeptor-immunpositiver Neurone im SP und IPS. Insgesamt zeigt sich eine konstante Verteilung in der caudo-rostralen Ausdehnung des SP und IPS. Links-rechts Unterschiede sind nicht festzustellen.
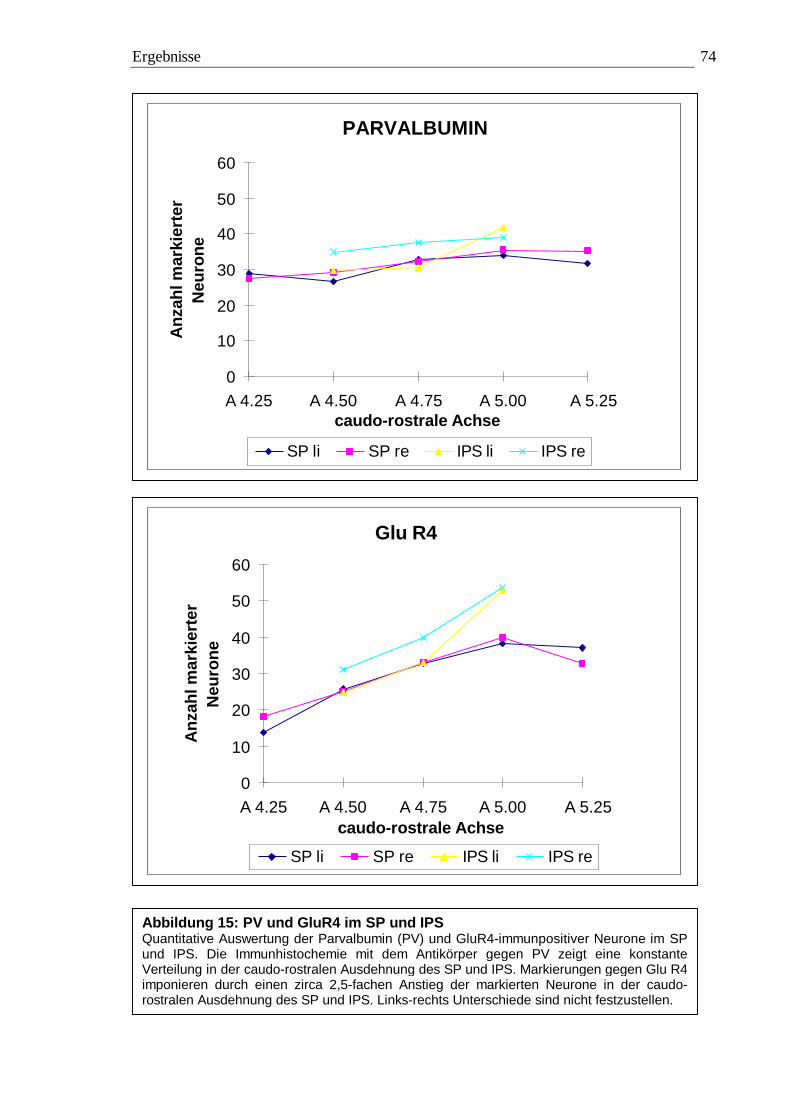
Ergebnisse
74
PARVALBUMIN
0
10
20
30
40
50
60
A 4.25 A 4.50 A 4.75 A 5.00 A 5.25caudo-rostrale Achse
Anz
ahl m
arki
erte
r N
euro
ne
SP li SP re IPS li IPS re
Glu R4
0
10
20
30
40
50
60
A 4.25 A 4.50 A 4.75 A 5.00 A 5.25caudo-rostrale Achse
Anz
ahl m
arki
erte
r N
euro
ne
SP li SP re IPS li IPS re
Abbildung 15: PV und GluR4 im SP und IPS Quantitative Auswertung der Parvalbumin (PV) und GluR4-immunpositiver Neurone im SP und IPS. Die Immunhistochemie mit dem Antikörper gegen PV zeigt eine konstante Verteilung in der caudo-rostralen Ausdehnung des SP und IPS. Markierungen gegen Glu R4 imponieren durch einen zirca 2,5-fachen Anstieg der markierten Neurone in der caudo-rostralen Ausdehnung des SP und IPS. Links-rechts Unterschiede sind nicht festzustellen.

Ergebnisse
75
Im Detail war die Anzahl GAD-markierter Neuronen im Nucleus subpraetectalis
von caudal nach rostral gleichmäßig verteilt (Abbildung 14). Auch die GABAA-
Rezeptor-positiven und Parvalbumin-positiven Neuronen wiesen im Nucleus
subpraetectalis eine konstante Verteilung auf der caudo-rostralen Achse auf, mit
dem Unterschied, daß die absolute Zellzahl der Parvalbumin-positiven Neuronen
etwas höher lag, als die GAD und den GABAA-Rezeptor exprimierenden
Nervenzellen (Abbildungen 14 und 15). In der statistischen Auswertung dieser
Daten mittels einer „Zwei-Wege ANOVA“ mit anschließender „post-hoc
Untersuchung (Scheffé-Test)“ zeigte sich, daß sowohl GAD und GABAA-Rezeptor,
als auch Parvalbumin exprimierende Neuronen keine signifikanten Unterschiede in
der Verteilung über die caudo-rostrale Achse des Nucleus subpraetectalis, als
auch zwischen beiden Hemisphären des Taubengehirns aufwiesen (GAD: n=5,
p? 0.05; GABAA-Rezeptor: n=5, p? 0.05; Parvalbumin: n=6; p? 0.05).
Die quantitativen Auswertungen Glu R4 markierter Neuronen ergab, daß die
Anzahl immunpositiver Neuronen auf der Ebene A5.25 um zirka den Faktor 2,5
höher lag als auf der Ebene A4.25 (Abbildung 15). In der statistischen Analyse
dieser Daten wurde deutlich, daß entlang der caudo-rostralen Ausdehnung dieses
Kernes die Anzahl der Glu R4 synthetisierenden Zellen signifikant anstieg (n=6,
p<0.01), während keine signifikanten links-rechts Unterschiede gemessen wurden
(p>0.05).
Abbildung 16 faßt den Anteil immunpositiver Neuronen gegenüber den
Zellzählungen in Kresylviolett-Präparaten in Prozent für den Nucleus
subpraetectalis zusammen. Dabei wurde für GAD und den GABAA-Rezeptor eine
konstante Expression dieser Proteine im caudalen und rostralen Nucleus
subpraetectalis deutlich. Die gesamte Verteilung GAD-postiver Zellen im Nucleus
subpraetectalis variierte zwischen 65% im rostralen Bereich und 76% im caudalen
Nucleus subpraetectalis (Abbildung 16). Der prozentuale Anteil der mit den
GABAA-Rezeptor markierter Neuronen lag zwischen 58-83%, wobei die Werte von
83%, 74% und 75% in den Hirnebenen A4.25, A4.75 und A5.25 beobachtet
wurden, und die Ebenen A4.50 und A.5.00 im Nucleus subpraetectalis mit 66%
und 58% anteilig geringer markiert waren. Auch bei Parvalbumin zeigte sich die
gleichmäßige Markierung von caudal nach rostral mit prozentualen Werten
zwischen 88% und 97%.
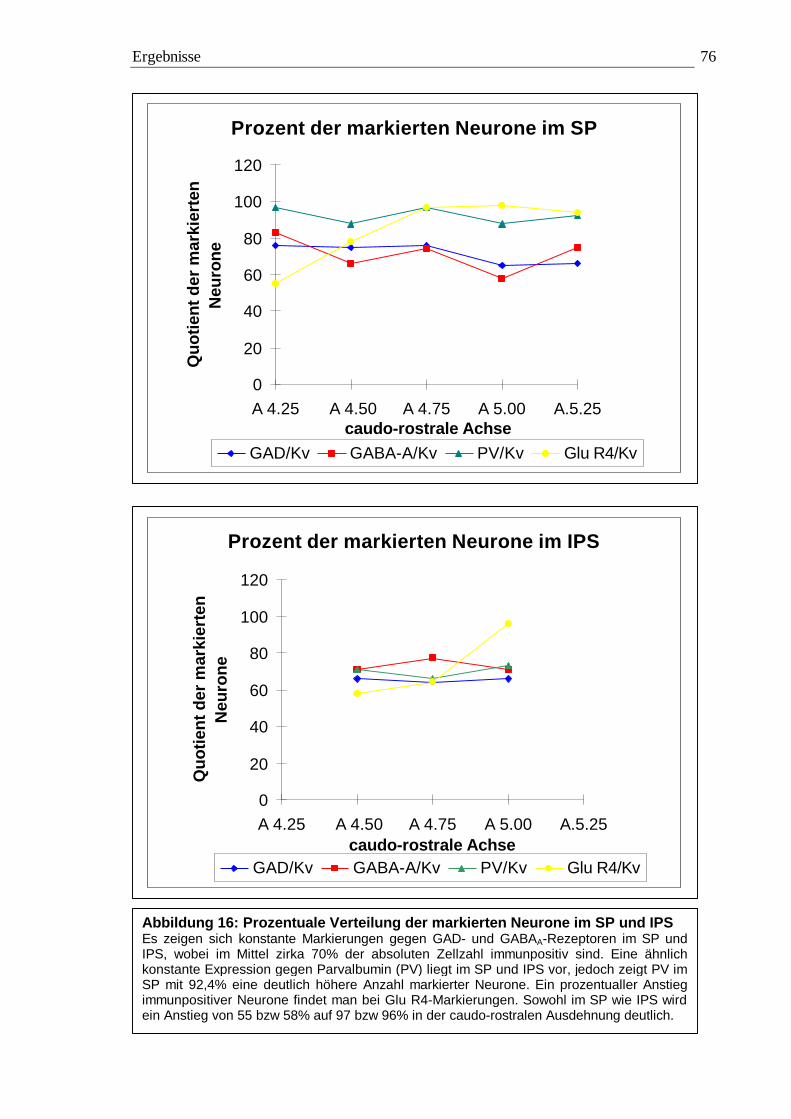
Ergebnisse
76
Abbildung 16: Prozentuale Verteilung der markierten Neurone im SP und IPS Es zeigen sich konstante Markierungen gegen GAD- und GABAA-Rezeptoren im SP und IPS, wobei im Mittel zirka 70% der absoluten Zellzahl immunpositiv sind. Eine ähnlich konstante Expression gegen Parvalbumin (PV) liegt im SP und IPS vor, jedoch zeigt PV im SP mit 92,4% eine deutlich höhere Anzahl markierter Neurone. Ein prozentualler Anstieg immunpositiver Neurone findet man bei Glu R4-Markierungen. Sowohl im SP wie IPS wird ein Anstieg von 55 bzw 58% auf 97 bzw 96% in der caudo-rostralen Ausdehnung deutlich.
Prozent der markierten Neurone im SP
0
20
40
60
80
100
120
A 4.25 A 4.50 A 4.75 A 5.00 A.5.25caudo-rostrale Achse
Quo
tient
der
mar
kier
ten
Neu
rone
GAD/Kv GABA-A/Kv PV/Kv Glu R4/Kv
Prozent der markierten Neurone im IPS
0
20
40
60
80
100
120
A 4.25 A 4.50 A 4.75 A 5.00 A.5.25caudo-rostrale Achse
Quo
tient
der
mar
kier
ten
Neu
rone
GAD/Kv GABA-A/Kv PV/Kv Glu R4/Kv

Ergebnisse
77
Ein deutlicher prozentualer Anstieg immunpositiver Neuronen im Nucleus
subpraetectalis wurde für die Rezeptoruntereinheit Glu R4 gezeigt, mit einem
Anstieg von 55% auf 97% zwischen A4.25 und A4.75. Weiter posterior (bis A5.25)
blieb der prozentuale Anteil Glu R4 markierter Neuronen konstant. Diese
Ergebnisse zeigen eine konstante Verteilung von GAD- und GABAA-Rezeptor-
positiver Neurone, im Mittel 71,6% und 71,2%, und einen hohen Prozentsatz von
zirka 84,4% Glu R4- und 92,4% Parvalbumin-positiver Neurone im Nucleus
subpraetectalis.
3.2.3.2 Quantitative Auswertung im Nucleus interstitio -praetecto-
subpraetectalis
Die quantitative Auswertung im Nucleus interstitio-praetecto-subpraetectalis
offenbarte viele Ähnlichkeiten in dem Expressionsmuster von GAD, dem GABAA-
Rezeptor, Parvalbumin und Glu R4 zu dem benachbarten Nucleus
subpraetectalis.
Die Antiköper gegen GAD, den GABAA-Rezeptor und das Calcium-bindende
Protein Parvalbumin markierten Neuronen ohne signifikante Unterschiede in der
caudo-rostralen Ausdehnung zwischen A4.50-A5.00 des Nucleus interstitio-
praetecto-subpraetectalis (Abbildung 16), sowie zwischen den beiden
Hemisphären (GAD: n=5, p? 0.05; GABAA-Rezeptor: n=5, p? 0.05; Parvalbumin:
n=6; p? 0.05). Hingegen nahm die Anzahl Glu R4-positiver Neuronen zwischen
A4.50 und A5.00 signifikant zu (n=6, p<0.01) (Abbildung 16)., ohne jedoch, daß
signifikante links-rechts Unterschiede offensichtlich wurden.
Bei der Bestimmung der prozentualen Verteilung zeigten sich keine Unterschiede
zwischen der Expression von GAD, GABAA-Rezeptor und Parvalbumin
exprimierender Neuronen im Nucleus interstitio-praetecto-subpraetectalis. Der
Anteil GAD markierter Neuronen variierte zwischen 64-66%, der Anteil an GABAA-
Rezeptor-positiven Neuronen lag zwischen 71-77%, und bei den Parvalbumin-
positiven Neuronen lag die Varianz zwischen 66-73%. Demgegenüber wurde bei
der Darstellung der Glutamatrezeptoruntereinheit Glu R4 zwischen A4.50 und
A5.00 ein Anstieg von 58% auf 96% deutlich, so daß im rostralen Nucleus
interstitio-praetecto-subpraetectalis nahezu alle Neuronen eine Glu R4 Markierung
aufwiesen.

Diskussion
78
4 Methodendiskussion
4.1 Injektionen von BDA und CtB
In der vorliegenden Untersuchung wurden die Konnektivitäten des Nucleus
subpraetectalis (SP) und des Nucleus interstitio-praetecto-subpraetectalis (IPS)
insbesondere zum tectofugalen System der Taube Columba livia untersucht.
Darüber hinaus sollten mit Hilfe immunhistochemischer Studien mögliche
exzitatorische und inhibitorische Signalwege dieser praetectalen Kerne erfaßt
werden.
Die Verbindungen des SP und IPS zu weiteren Hirnstrukturen wurden dabei durch
Injektionen neuronaler Tracer wie Biotin-Dextran-Amin (BDA) sowie die
ß-Untereinheit des Choleratoxin (CtB) in den SP oder IPS erfaßt. Bei beiden
Tracern handelt es sich um Substanzen, die nach Applikation in das
entsprechende nervale Gewebe, von den dort gelegenen Nervenzellen
phagocytiert werden. Während BDA anschließend zumeist anterograd verbundene
Hirnstrukturen markiert (Brandt & Apkarian, 1992), wurde CtB als vornehmlich
retrograder Tracer eingesetzt (Angelucci et al., 1996; Hellmann & Güntürkün,
1999). In beiden Richtungen, anterograd und retrograd, handelt es sich vermutlich
um aktiven Transport, bei dem wahrscheinlich zunächst eine vesikuläre
Verpackung der injizierten Substanz erfolgt, welche dann von spezifischen
Transportermolekülen wie Kinesin (Hirokawa, 1998) transportiert wird. Als
anterograder Transport wird hierbei die Verfrachtung der injizierten Substanz vom
Somata zur Nervenendigung bezeichnet, während retrograder Transport die
Transportrichtung von der Nervenendigung, der Synapse zum Zellsomata
beschreibt. Mit Hilfe der beiden Tracer BDA und CtB konnten somit nach Injektion
in den SP sowohl afferente als auch efferente Verbindungen dieser praetectalen
Kerne erfaßt werden.
Die Zeit, bis der Tracer die Nervenendigung beziehungsweise das Zellsomata
erreicht, war dabei abhängig von der Transportstrecke sowie vom
Molekulargewicht der Tracer. BDA ist mit einem Molekulargewicht von 10000Da
ein großer Tracer, dessen Membranpermeabilität am Injektionsort durch Zusatz
von DMSO verbessert wurde. CtB war hingegen auch ohne weiteren Zusatz gut
membranpermeabel, und wurde von den Somata und Nervenendigungen direkt
aufgenommen. Die bedeutenden Unterschiede der beiden Tracer sind, neben

Diskussion
79
ihrer bevorzugt anterograden oder retrograden Transportrichtung, daß CtB eine
detaillierte Darstellung von Axonen in ihrer gesamten Länge mit beinahe Golgi-
ähnlicher Auflösung von Terminalien ermöglicht, während sich BDA auf eine
Markierung der Somata im verbundenen Kerngebiet beschränkt (Angelucci et
al.,1996). In der vorliegenden Arbeit zeigte sich die deutlich erhöhte
Membranpermeabilität und der gesteigerte Transport von CtB unter anderem
darin, daß am Injektionsort keine Tracerrückstände von CtB drei Tage nach der
Injektion zu erkennen waren, während der BDA Injektionsort auch nach sieben
Tagen Überlebenszeit gut zu erkennen war.
Diese offensichtlichen Unterschiede der beiden Tracer wurden in der
gegenwärtigen Untersuchung genutzt, um 3 bis 7 Tage nach der Tracerinjektion
die afferenten und efferenten Konnektivitäten des SP und IPS innerhalb des
tectofugalen Systems und zum Teil darüber hinaus nach entsprechender
histochemischer Aufbereitung darzustellen.
4.2 Immunhistochemie
Mittels der Methode der Immunhistochemie wird die Expression von Proteinen
durch spezifische, gegen diese gerichtete Antikörper in Zellen nachgewiesen. Bei
dem Einsatz geeigneter Antikörper kann hierbei im Idealfall eine genaue
Lokalisation der Proteine in den Zellen erfolgen. In Nervenzellen ist somit zum
Beispiel eine genaue Lokalisation von Rezeptorproteinen im Somata oder den
Dendriten möglich. Die eingesetzten Antikörper gegen den GABAA-Rezeptor und
den Glu R4-Rezeptor, sowie gegen GAD und Parvalbumin zeigten gleichermaßen
somatische Markierungen, wobei der GABAA-Rezeptor, GAD und Parvalbumin
darüber hinaus auch im Neuropil nachgewiesen wurden. Immunhistochemische
Anfärbungen des Somata deuten darauf hin, das die entsprechenden Proteine im
Zellsomata exprimiert und vermutlich auch lokalisiert sind. Für den GABAA-
Rezeptor und Glu R4 kennzeichnet dies das Vorhandensein axo-somatischer,
membrangebundener Rezeptoren in diesen Nervenzellen. Auch die beiden
Proteine GAD und Parvalbumin liegen demnach massiv, vermutlich cytosolisch in
den Somata entsprechend immunpositiver Neuronen vor. Immunmarkierungen
des Neuropil zeigen außerdem die Lokalisation der Proteine GAD, Parvalbumin,
und des GABAA-Rezeptors in den Dendriten der entsprechenden Nervenzellen an.
Während der GABAA-Rezeptor vermutlich auch hier membrangebunden an den

Diskussion
80
Synapsen dieser Zellfortsätze vorliegt, so ist für GAD und Parvalbumin ein
vermutlich cytoplasmatisches Vorkommen in diesen Zellfortsätzen wahrscheinlich.
Gleichwohl sind diese Aussagen nur aus der Funktion der Proteine abzuleiten
(siehe Diskussion der biochemischen Signalwege), da keine ultrastrukturellen
Daten über die Lokalisation der Proteine vorliegen.
Die Spezifität der eingesetzten Antikörper wurde seitens der Anbieter und in
diversen vorangegangenen Untersuchungen überprüft. Auch in
Kontrollexperimenten der vorliegenden Arbeit zeigte sich, daß ein Ersatz des
primären Antikörpers durch ein unspezifisches Serum zu keinerlei Markierungen
im aufbereiteten Hirngewebe der Taube führte.

Diskussion
81
5 Verschaltungsmuster des Nucleus subpraetectalis
Nach Injektion der beiden Tracer BDA und CtB in den Nucleus subpraetectalis
stellten sich vier unterschiedliche Applikationsorte dar. Injiziiert wurde in den
ventralen, ventro-lateralen, in den lateralen sowie dorsalen Nucleus
subpraetectalis (SP). Die im folgenden kurz skizzierten Konnektivitäten des SP
beziehen sich auf das mesencephale Tectum opticum (TO), dem zu den
praetectalen Kernen zugehörige Nucleus interstitio-praetecto-subpraetectalis (IPS)
und den Nucleus praetectalis (PT) sowie auf den diencephalen Nucleus rotundus
(RT). Abhängig vom Injektionsort im SP zeigte sich eine differente topographische
Markierung in den einzelnen Kernen, wobei Unterschiede in der Dichte und
Lokalisation der markierten Strukturen, zum Beispiel in der Anzahl und der
Lokalisation der markierten Somata oder Fasern, deutlich wurden.
Injektionen in den ventralen SP führten im IPS zu einer deutlichen und
gleichmäßigen Anfärbung des Neuropil, während somatische Markierungen nicht
zu erkennen waren. Hingegen waren im TO in der Schicht 13 gelegene Somata
sowie das Neuropil dieser Schicht gleichmäßig in der gesamten dorso-ventralen
Ausdehnung, ohne Unterschiede in der rostro-caudalen Achse gekennzeichnet.
Injektionen von CtB oder BDA in den ventralen SP markierten im RT
ausschließlich das Neuropil. Hierbei zeigte sich ein Markierungsmuster, nach
welchem der ventrale SP auf den gesamten rostralen RT projiziert, während
caudal nur der laterale RT vom ventralen SP innerviert wird. Im PT, waren nach
Injektionen in den ventralen SP Faserterminalien markiert, während vemutlich
einziehende Fasern nur in den äußeren Anteilen dieses Kernes zu erkennen
waren.
Nach Applikation der Tracer in den ventro-lateralen SP stellten sich ähnliche
Projektionen dar. Im IPS zeigten sich vereinzelt somatische Markierungen und
eine gleichmäßige Anfärbung des Neuropil. Das TO war dagegen durch intensive
somatische Markierungen in der Schicht 13 gekennzeichnet, die von intensiven
Neuropilfärbungen in dieser multipolaren tectalen Schicht begleitet waren. In einer
subjektiven Betrachtung zeigte sich hierbei eine qualitative Dichtezunahme dieser
Markierungen im ventralen TO. Injektionen in den ventro-lateralen SP führten im
RT zu Anfärbungen des Neuropil, wobei auch hier caudal primär der laterale RT
markiert war, während weiter rostral eine Ausdehnung dieser Neuropil-Markierung

Diskussion
82
auf den gesamten RT zu erkennen war. Im PT wurden wiederum
Neuropilanfärbungen in den äußeren Bereichen des Kernes nach Injektionen der
Tracer in den ventro-lateralen SP deutlich.
Wurde der Tracer in den lateralen SP appliziert, so war im rostralen IPS der
gesamte Kern durch markierte Faser gekennzeichnet. Im mesencephalen TO fand
sich eine deutliche Anfärbung der Somata und des Neuropil in der Schicht 13,
wobei eine intensivere und subjektiv dichtere Markierung der Schicht 13 des
ventralen TO gegenüber dem dorsalen TO deutlich wurde. Ebenso wie nach
Tracerinjektionen in den ventralen und ventro-lateralen SP, zeigten sich auch nach
CtB- oder BDA-Injektionen in den lateralen SP primär laterale Faser- und
Terminalienmarkierungen im caudalen RT, die sich weiter rostral zum medialen
RT ausbreiteten. Im rostralen RT stellten sich im gesamten Kern
Fasermarkierungen nach Injektionen in den lateralen SP dar, wobei keinerlei
somatische Markierungen zu erkennen waren. Im PT hingegen wurden
Terminalien im gesamten Kern sowie Faser im äußeren Bereich des Kernes nach
Injektionen in den lateralen SP dargestellt.
Nachdem der Tracer in den dorsalen SP injiziiert wurde, konnten Projektionen
dieses Bereiches auf den IPS nachgewiesen werden, wobei vereinzelt Somata,
zumeist aber das Neuropil in diesem Kern dargestellt wurde. Im TO wurden
Somata und Fasern der Schicht 13 gleichmäßig in der dorso-ventralen, und rostro-
caudalen Ausdehnung markiert. Der RT war gekennzeichnet durch eine
Fasermarkierung, die sich caudal auf den lateralen RT beschränkte, im zentralen
RT keinerlei Markierungen zeigte, um im rostralen RT Neuropil im gesamten Kern
zu markieren. Im PT zeigte sich nach Tracerinjektion in den dorsalen SP eine
Anfärbung vereinzelter Somata, konträr zu den Ergebnissen nach Injektionen in
den ventralen, ventro-lateralen oder lateralen SP. Außerdem waren aber auch hier
deutliche Markierungen des Neuropil im gesamten Kern zu verzeichnen, wobei
aber auch hier die lateralen Abschnitte intensiver markiert waren.
Zusammenfassend stellte man fest, daß je nach Farbstoffinjektion in den SP ein
ähnliches Markierungsmuster in den beschriebenen Kernen vorliegt. Der IPS
zeigte deutliche Neuropilanfärbung und teilweise vereinzelt, nach ventro-lateraler
und dorsaler Injektion der Tracer in den SP markierte Somata. Im TO beobachtete
man unabhängig von der Tracerapplikation im SP eine Anfärbung der Somata und
des Neuropil in Schicht 13, jedoch bestand nach ventro-lateraler und lateraler

Diskussion
83
Farbstoffinjektion in den SP ein von ventral nach dorsal abnehmender
Zellzahlgradient in der Schicht 13 des TO. Im RT fand sich nach jeder
Tracerinjektion eine ähnliche Anfärbung der Faser. In rostralen Abschnitten war
der gesamte Kern markiert und im caudalen Anteil beschränkte sich die
Fasermarkierung auf den lateralen RT. Zuletzt sei noch der PT erwähnt, der
hauptsächlich durch eine Neuropilanfärbung in den äußeren Bereichen auffiel.
5.1 Tectum opticum - Nucleus subpraetectalis
Die Verbindung des mesencephalen TO zum SP/IPS-Komplex wurde durch
unterschiedliche wissenschaftliche Gruppen bei Hühnchen, Zebrafinken und
Tauben untersucht, wobei keine topographische Organisation innerhalb der
Projektion der tectalen multipolaren Schicht 13 auf den SP/IPS erkannt wurde
(Karten & Revzin, 1966; Hunt & Künzle, 1976; Bischof & Niemann, 1990).
Retrograde Markierungen nach CtB-Injektionen in den SP der vorliegenden Arbeit
bestätigen diese Ergebnisse, wobei sich eine massive Markierung der Schicht 13
Neuronen zeigte, begleitet von einem dorso-ventral zunehmenden
Zellzahlgradienten in dieser Schicht. Vermutlich erhält der SP einen stärkeren
Input aus dem ventralen TO gegenüber dem dorsalen TO. Tectale
Tracerinjektionen zeigten, daß der Nucleus rotundus den größten Teil der tectalen
Eingänge von ventralen Schicht 13 Neuronen erhält (Hellmann & Güntürkün,
1999). Die Autoren postulieren eine Gesichtsfeld abhängige Projektion, da im
ventralen TO die visuelle Information des frontalen Gesichtsfeldes repräsentiert ist
(Remy & Güntürkün, 1991). Möglicherweise trägt der SP/IPS-Komplex zu
größeren Teilen zur Informationsverarbeitung in diesem Gesichtsfeld bei. Jedoch
besteht auch die Möglichkeit einer analogen gesichtsfeldabhängigen
Repräsentation in beiden Projektionssystemen, daß heißt tecto-rotundal, sowie
tecto-praetecto-rotundal. Damit bestände die Funktionalität des zusätzlichen
Projektionssystems nicht in der Informationsergänzung, sondern vielmehr in der
unterschiedlichen Informationsverarbeitung. Diese könnte durch einen „direkten“,
schnellen tecto-rotundalen Weg und einer „indirekten“, langsamen tecto-praetecto-
rotundalen Projektion gekennzeichnet sein.
Mpodozis und Mitarbeiter (1996) stellten spärliche Projektionen vom TO zum
kontralateralen SP/IPS-Komplex bei Tauben fest. Ähnliche Daten beschrieben
Bischof und Niemann (1990) bei Zebrafinken. Sie zeigten daß der SP/IPS-

Diskussion
84
Komplex bilaterale tectale Eingänge erhält. Nach Injektion der Tracer BDA und
CtB in den SP fanden sich in der vorliegenden Arbeit jedoch keine Markierungen
im kontralateralen TO. Eine Ursache dafür könnte eine zu geringe
Diffusionskapazität der beiden Tracer sein. Vielleicht waren die Überlebenszeiten
der Tauben von drei bis sieben Tagen zu gering gewählt, so daß der axonale
Transport dieser Substanzen bis zum kontralateralen TO nicht abgeschlossen
war. Es ist aber auch denkbar, daß der mediale SP kontralateralen tectalen Input
erhält, ein Bereich des SP, der in der gegenwärtigen Arbeit von den
Tracerapplikationen unberührt geblieben ist.
5.2 Nucleus subpraetectalis – Nucleus rotundus
Benowitz und Karten (1976) beschreiben die Projektion zwischen SP/IPS-Komplex
und dem diencephalen RT in der Taube, wonach der RT vermutlich inhibitorischen
Input aus diesen praetectalen Kernen erhält. Efferenzen des SP/IPS-Komplexes
beschränken sich gemäß den Autoren jedoch auf den ventralen und antero-
ventro-lateralen Bereich des RT. Spätere Arbeiten von Deng und Rogers (1998)
und Mpodozis und Mitarbeiter (1996) zeigen bei Hühnchen und Tauben, daß der
SP/IPS-Komplex auf den gesamten RT projiziert. Deng und Rogers konkretisieren
die SP Projektion auf den RT; demnach projiziert der dorsale SP zum dorsalen
RT, der ventrale SP zum ventralen RT sowie der mittlere Bereich des SP auf den
zentralen RT.
In den gegenwärtigen Untersuchungen bei der Taube wurden nach Injektionen in
den SP in caudalen Hirnabschnitten der laterale RT, und weiter rostral der
gesamte RT markiert. Gemeinsam ist allen beschriebenen Untersuchungen, daß
der SP/IPS-Komplex nur ipsilateral auf den RT projiziert. Tömböl und Mitarbeiter
(1994) untersuchten die Projektionen vom SP und PV auf den RT bei Hühnchen,
wobei sie GABAerge Strukturen an den markierten Nervenendigungen im RT
nachwiesen. Neben diesen Tracinguntersuchungen zeigten auch immunologische
und Läsionsstudien von Mpodozis (1996) nach Läsion des SP eine Abnahme der
GABAergen Neuropilmarkierung im RT.
Zusammenfassend bestätigen diese Daten den Einfluß des SP/IPS-Systems auf
das tectofugale System und damit seine wichtige Rolle bei der Verarbeitung
visueller Informationen im RT.

Diskussion
85
5.3 Nucleus subpraetectalis -Nucleus triangularis
Der Nucleus triangularis (T) liegt als dorsale Kappe dem RT an. Bislang gibt es
aber kaum Erkenntnisse über die Konnektivitäten dieses Kernes, als auch über
mögliche physiologische Relevanzen, zum Beispiel bei der Verarbeitung visueller
Informationen. In einer Konnektivitätsstudie wurde gezeigt, daß der T keine
direkten Verbindungen mit dem SP besitzt (Benowitz & Karten, 1976; Hellmann &
Güntürkün, 2001), während jedoch somatische Markierungen im IPS und nTT
(Nucleus tectothalamicus) eine Projektionen vom IPS zum T anzeigen (Deng &
Rogers, 1998; Hellmann & Güntürkün, 2001).
In der vorliegenden Arbeit blieb der T nach SP-Injektionen von jeglicher
Markierung ausgespart. Somit zeigte sich auch mit den Tracern BDA und CtB, daß
keine direkten Verbindungen zwischen dem T und dem SP besteht.
5.4 Nucleus subpraetectalis - Nucleus praetectalis
Der PT wurde in bisherigen Untersuchungen nur sehr spärlich beschrieben. Eine
Untersuchung von Gamlin und Mitarbeitern (1996) bei Tauben verweist auf
Konnektivitäten zwischen dem PT und dem TO, wobei Neuronen der Schicht 5b
auf den PT projizieren. Nach Tracerinjektionen in den SP zeigten sich in der
gegenwärtigen Arbeit deutliche Markierungen des Neuropil, sowie sehr vereinzelte
somatische Markierungen im PT. Da gleichzeitig keine Markierungen in der
Schicht 5b des TO erkannt werden konnten, handelt es sich möglicherweise nicht
um retrograde Projektionen des TO zum PT, sondern um eine direkte SP – PT
Projektion.
Hellmann und Güntürkün (2001) zeigten jedoch, daß Neuronen der Schicht 13
teilweise auf den PT projizieren. Daher ist es in dieser Arbeit nicht
auszuschließen, daß die PT Markierungen durch retrograde TO Projektion
hervorgerufen wurden.

Diskussion
86
6 Biochemische Organisation des SP/IPS-Komplexes
Mit Hilfe immunhistochemischer Untersuchungen konnte in der vorliegenden
Arbeit die bislang weitgehend unbekannte biochemische Organisation der
praetectalen Kerne SP und IPS bei der Taube näher charakterisiert werden. Die
Immunmarkierungen gegen den GABAA- und den GluR4 Rezeptor sind hierbei ein
Hinweis auf inhibitorische und exzitatorische afferente Signalverarbeitungswege.
Die somatische Verteilung der beiden Proteine GABAA und GluR4 kennzeichnet
dabei vermutlich axo-somatische inhibitorische und exzitatorische Rezeptoren,
während das GABAA-positive Neuropil-Signal wahrscheinlich Ausdruck
dendritischer GABAA-Rezeptoren ist.
Die massiven somatischen und im Neuropil lokalisierten GAD-Signale der
vorliegenden Arbeit sind ein deutlicher Hinweis auf die GABA Synthese in
Neuronen des SP und IPS bei der Taube. Eine hohe Dichte GABAerger Neuronen
wurde auch von Domenici und Mitarbeitern (1988) in diesen Kernen des
Taubenhirns erkannt. Die Autoren stellten fest, daß das somatische GABA-Signal
gegenüber den Neuropil-Markierungen hervortrat, wodurch sie auf eine
prominente GABAerge Projektion aus den beiden Kernen zu anderen neuronalen
Kerngebieten rückschlossen. Gleichwohl sind auf Grund der in der vorliegenden
Arbeit vorgestellten Markierungen der Axone und Dendriten in diesen praetectalen
Kernen auch intrinsische GABAerge Signalwege denkbar, die unter
Gliederungspunkt 7 in ein Modellsystem integriert werden. Ein mögliches
Terminationsgebiet einer solchen inhibitorischen Projektion ist der Nucleus
rotundus. In ultrastrukturellen Untersuchungen bei Hühnchen wurden Gruppen
von axo-somatischen und axo-dendritischen Terminalien im Nucleus rotundus
identifiziert, deren Synapsen symmetrisch aufgebaut waren und abgeflachte
Vesikeln enthielten, die GABA-positiv waren (Ngo et al., 1992). Die Autoren dieser
Studie spekulierten bereits, daß diese Terminalien von Axonen aus dem Nucleus
subpraetectalis stammen. In einer kombinierten ultrastrukturellen Tracing und
immunhistochemischen Studie wurde diese Vermutung bei Hühnchen bestätigt,
wonach die Projektionen des SP und auch des PV auf den Nucleus rotundus
GABA-erg sind (Tömböl et al., 1994).
Die somatische Expression von Parvalbumin, ebenfalls begleitet von
Neuropilmarkierungen korreliert vermutlich mit den GAD-positiven Neuronen im

Diskussion
87
SP/IPS-Komplex. Im Neocortex sowie im Hippocampus wurde das Calcium-
bindende Protein Parvalbumin kolokalisiert mit GABAergen Neuronen detektiert,
Neuronen welche eine hohe Feuerfrequenz kennzeichnet (Celio, 1986; Freund,
1989). Gleichwohl ist das Vorkommen von Parvalbumin nicht spezifisch für
einzelne Transmitter, wie Braun und Mitarbeiter (1988) für den Nucleus rotundus
zeigen konnten. In diesem diencephalen Kern wurden keine GABAergen
Neuronen, wohl aber Parvalbumin-positive Neuronen lokalisiert. Bei der Taube
zeigte sich im Nucleus rotundus eine Kolokalisation von Glu R4 und Parvalbumin
(Theiss et al., 1998).
Die aufgezeigten Daten machen deutlich, daß Glutamat vermutlich als
exzitatorischer Transmitter, GABA wahrscheinlich als inhibitorischer
Neurotransmitter auf Neuronen des SP und IPS fungiert. Auf Grund der massiven
somatischen GAD-Expression und den Daten anderer Studien zufolge ist GABA
darüber hinaus als Neurotransmitter der efferenten Projektionen dieser Kerne
anzunehmen. Ein Modell über die möglichen Signalwege des SP/IPS-Komplexes,
insbesondere im Bezug zur Signalverarbeitung im tectofugalen System der Taube,
wird im folgenden Gliederungspunkt 7 diskutiert.

Diskussion
88
7 Integration des SP/IPS-Komplexes in das tectofugale System
Bei den Mammaliern erfolgt die Diskrimination und Verarbeitung visueller
Informationen weitgehend im retino-genikulocorticalen System, während bei
Vögeln und Reptilien eine solche visuelle Signalfortleitung größtenteils in den
Kerngebieten der tectofugalen Sehbahn geleistet wird. Bei den Vögeln und
Reptilien nimmt die Projektion von der Retina zum Corpus geniculatum und weiter
zum visuellen Wulst im Vorderhirn, auch als thalamofugale Sehbahn bezeichnet
und dem retino-genikulocorticalen System der Mammalier homolog, nur einen
kleinen Teil der retinalen Projektionen ein. Hingegen terminieren etwa 90%
retinaler Axone im mesencephalen TO, um von dort bilateral über den
diencephalen RT zum im Vorderhirn gelegenen Ectostriatum zu projizieren (Karten
& Hodos, 1970). Der RT fungiert somit als thalamisches Relaycenter bei der
Weiterleitung visueller Informationen zum Telencephalon (Engelage & Bischof,
1989). Innerhalb der tectofugalen Sehbahn erhält er seine Afferenzen bilateral aus
der tectalen Schicht 13 (Karten & Revzin, 1966), wobei Neuronen in
unterschiedlichen Lagen innerhalb dieser Schicht 13 auf distinkte Bereiche des RT
projizieren (Benowitz & Karten, 1976; Luksch et al., 1998; Hellmann et al., 1999).
In diversen anatomischen Untersuchungen und Verhaltensstudien wurde gezeigt,
daß für die visuelle Signalverarbeitung bei Vögeln und Reptilien maßgeblich die
weitere Projektion vom RT zum Ectostriatum verantwortlich ist (Hodos & Karten,
1966, 1970; Reiner & Powers, 1980; Shimizu & Karten, 1993). Im Vergleich zu
den Mammaliern ordnen Shimizu und Karten (1993) den RT als homolog zur
inferioren caudalen Pulvinar ein, einem bei Mammaliern multimodal arrangierten
Kerngebiet, daß möglicherweise auch an der visuellen Signalverarbeitung beteiligt
ist (Raczkowski & Diamond, 1980).
Anatomische, biochemische und physiologische Untersuchungen teilen den RT
der Vögel in diverse Subregionen ein. So zeigten Benowitz und Karten (1976)
sowie Nixdorf und Bischof (1982) anhand von Tracinguntersuchungen, daß der
posteriore RT Afferenzen der tiefen tectalen Schicht 13 erhält, während die
multipolaren Neuronen der oberen Schicht 13 auf den dorsalen und anterio-
medialen RT projizieren. Darüber hinaus terminieren Axone von in
unterschiedlichen Höhen gelegenen tetalen Schicht 13 Neuronen im gesamten

Diskussion
89
ventralen RT. In einer weiteren Studie wurde der RT mittels des biochemischen
Nachweises von Acetylcholin Esterase (AChE) unterteilt (Martinez-de-la Torre,
1990). Hierbei konnte AChE nicht im postero-lateralen, im postero-medialen, im
antero-medialen und rostralen RT nachgewiesen werden, während der
intermediale RT AchE-positiv war. Immunhistochemisch wurde gezeigt, daß die
Expression von Glutamatrezeptoren im RT ebenfalls zwischen dorsalen und
ventralen rotundalen Bereichen variiert (Theiss et al., 1998). Auch
elektrophysiologisch war eine Substrukturierungen des RT möglich (Granda &
Yazulla, 1971; Revzin, 1979; Wang et al., 1993). Demnach verarbeitet der ventrale
RT vornehmlich bewegte Reize im 2-dimensionalen Raum, während antero-medial
Helligkeitsunterschiede und zum Teil bewegte Reize zu einer Erregung rotundaler
Neuronen führt. Im dorso-anterioren RT werden Farbinformationen empfangen
und weitergeleitet, während Bewegungen im 3-dimensionalen Raum im dorso-
posterioren und dorso-medialen RT gelegenen Neuronen stimulieren.
Diese Vielfalt von Substrukturen innerhalb des RT ist ein Indiz für die
Verschiedenartigkeit der Signalverarbeitung in diesem thalamischen Kern, dessen
Komplexität auch gegenwärtig noch nicht gänzlich erfaßt ist. Gleichwohl zeigen
auch die in der gegenwärtigen Untersuchung durchgeführten Tracingstudien mit
den beiden neuronalen Tracern BDA und CtB eine offensichtliche Strukturierung
der Projektionen des SP/IPS auf den RT. Deutliche Markierungen des Neuropil,
ohne das rotundale Somata markiert waren, zeigen die efferenten Projektionen
des SP auf den RT an. Diese Projektionen waren derart gegliedert, daß nach
ventraler oder ventro-lateraler Tracerapplikation caudal im diencephalen Kern
Neuropil im lateralen beziehungsweise ventro-lateralen RT markiert wurde,
während rostral in RT nach solchen Tracerapplikationen der gesamte Kern
Neuropilmarkierungen aufwies. Nach dem Tracing von BDA in den lateralen SP
wurden hingegen deutliche laterale Neuropilmarkierungen im caudalen RT
beobachtet, während auch hier weiter rostral der gesamte RT solche intensiven
Neuropilmakierungen aufwies. Die Injektionen von CtB in den dorsalen SP zeigten
dagegen ein Makierungsmuster von Neuropil im RT, wobei caudal Neuropil im
lateralen RT, und ventro-medial im gesamten RT markiert war.
Zusammenfassend lassen diese Daten eine Projektion des gesamten SP auf den
vornehmlich lateralen RT im caudalen Bereich des Kernes erkennen, während

Diskussion
90
rostral der gesamte RT vom SP innerviert wird. Eine topographische Organisation
dieser Projektionen vom dorsalen SP auf den dorsalen RT, beziehungsweise vom
ventralen SP auf den ventralen RT konnte somit nicht nachgewiesen werden. Für
eine sinnvolle Informationsverarbeitung innerhalb dieses Systems ist dies aber
auch keine Grundvoraussetzung, da bereits die Projektionen vom TO auf den RT
keiner retinotopen Ordnung folgen (Benowitz & Karten, 1976), sondern, wie oben
skizziert, vielmehr einer funktionellen Gliederung unterliegen. Unter solchen
funktionellen Gesichtspunkten projiziert der SP vornehmlich auf caudale rotundale
Bereiche, die an der Verarbeitung von Informationen über Bewegungen im
Gesichtsfeld beteiligt sind, während im rostralen RT der SP funktionelle Bereiche
innerviert, die Informationen über Farbe, Helligkeit und Bewegungen integrieren.
Hierbei stellt sich die Frage, welche Funktionen eine solche SP-Innervation des
RT bei der Verarbeitung visueller Informationen in diesem thalamischen Kern, der
eine wichtige, wenn nicht die wichtigste Integrationsstelle innerhalb der visuellen
Sehbahn auf dem Weg zum Vorderhin darstellt, ausübt. Um diese Frage zu
beantworten, mußten zunächst die Afferenzen des SP erfaßt werden. Mittels der
in der vorliegenden Arbeit durchgeführten Tracingstudien wurden neben den
Neuropilmarkierungen im RT auch sehr intensive somatische Markierungen von
Schicht 13 Neuronen im TO, begleitet von ebenfalls intensiven Markierungen des
Neuropil in dieser efferenten Schicht des TO, detektiert. Vereinzelt waren nach
Tracerinjektionen in den SP auch Neuronen in der gleichfalls efferenten tectalen
Schicht 10 histochemisch nachweisbar. Diese afferenten Konnektivitäten des SP
stimmen größtenteils mit Untersuchungen von Hunt und Künzle (1976a) überein.
In der gegenwärtigen Arbeit nicht bestätigt, von Hunt und Künzle aber postuliert,
projiziert das TO bilateral auf den SP. Auch bei Zebrafinken wurden solche ipsi-
und kontralateralen Projektion des TO auf den SP bestätigt (Bischof & Niemann,
1990). Möglicherweise liegt der Grund für die in der vorliegenden Arbeit
vornehmlich ipsilateralen Projektionen des TO auf den SP bei den verwendeten
Tracern BDA und CtB, während in den anderen beiden Untersuchungen andere
anterograde, in das TO injizierte Tracer zum Einsatz kamen. Gleichwohl dieser
Unterschiede konnte in der vorliegenden Arbeit mittels der eingesetzten Tracer
das Projektionsmuster des TO auf den SP noch einmal detailliert untersucht
werden, wobei ein abnehmender Dichtegradient somatischer Markierungen der
tectalen Schicht 13 von ventral nach dorsal offensichtlich wurde. Vermutlich ist

Diskussion
91
dieser Gradient aber nicht Ausdruck einer prozentual unterschiedlichen Projektion
zwischen ventralen und dorsalem TO auf den SP, sondern korreliert vielmehr mit
der von ventral nach dorsal abnehmenden Dichte der Neuronen in dieser Schicht,
wie in einer quantitativen Studie kresylviolett gefärbten Hirnschnitte deutlich wurde
(Theiss et al., 1998).
Offensichtlich gelangt bei Tauben die visuelle Information über zwei parallele
Wege zum RT. Der eine Informationsweg, die tectofugale Sehbahn, ist die direkte
Verbindung tectaler Schicht 13 Neuronen zum RT, während der Andere von
tectalen Neuronen der Schichten 10 und 13 zum SP, und dann weiter zum RT
verläuft. Tömböl und Mitarbeiter (1999) postulieren ein Modell, nach dem es sich
bei den ispilateralen Projektionen des TO auf den SP um Axonkollaterale der
ispilateralen Verbindungen zwischen TO und RT handelt.
Im Vergleich der hier vorgestellten Tracingdaten mit diversen
Konnektivitätsuntersuchungen zeigt sich, daß bei Tauben der SP/IPS-Komplex
vermutlich auf den ventralen (Benowitz & Karten, 1976), in der vorliegenden Arbeit
ventro-lateralen (caudal) bis gesamten RT (rostral) projiziert, während die weiteren
rotundalen Bereiche direkt vom TO innerviert werden. Bei Hühnchen hingegen
scheint der gesamte RT nebst Nucleus triangularis, der dorsalen Kappe dieses
diencephalen Kernes, vom SP/IPS-Komplex innerviert zu werden (Deng & Rogers,
1998). Gleichwohl konnten Hellmann und Güntürkün (2001) nach gezielten
dorsalen bis ventralen rotundalen Tracerapplikationen entsprechende dorsale bis
ventrale retrograde somatische Markierungen im SP erkennen. Für Hühner und
Taube gleichermaßen gilt, daß weder die Somata noch die Dendriten von
Neuronen des SP direkte retinale Eingänge erhalten, sondern die Afferenzen
dieses intrinsischen Kernes des tecto-thalamischen Traktes ausschließlich vom
TO kommen (Mpodozis et al., 1996).
Anhand der Daten der biochemischen Organisation des SP/IPS-Komplexes (siehe
Gliederungspunkt 6) und den beschriebenen Konnektivitäten, läßt sich ein Modell
über die Signalverarbeitung im tectofugalen System unter Einfluß der praetectalen
Kerne erstellen.

Diskussion
92
Gemäß dem in Abbildung 17 dargestellten Modell ist es denkbar, daß visuelle
Informationen von Neuronen der tectalen Schicht 13 zum einen direkt zu
rotundalen Neuronen geleitet werden, und parallel dazu zunächst zu Neuronen
des SP/IPS-Komplexes gelangen, um anschließend von diesen ausgehend den
RT zu innervieren. Auf Grund des in der vorliegenden Arbeit gezeigten GluR4-
Signals in den beiden praetectalen Kernen ist anzunehmen, daß diese tectalen
Efferenzen glutamaterge Eingänge erhalten. Die Innervation des RT vom SP und
IPS wiederum ist vermutlich inhibitorisch über den Neurotransmitter GABA. Wie
zuvor beschrieben exprimieren Neuronen dieser beiden praetectalen Kerne das
GABA synthetisierende Enzym GAD (eigene Befunde) sowie den Neurotransmitter
GABA (Domenici et al., 1988). In immunhistochemischen und ultrastrukturellen
Abbildung 17: Modell zur Signalverarbeitung im tecto-rotundalen System unter Einbeziehung des Nucleus subpraetectalis Neben der direkten (glutamatergen) Projektion tectaler Schicht 13 Neuronen zum Nucleus rotundus (RT), ist ein zweiter, polysynaptischer Informationsfluß über den Nucleus subpraetectalis (SP) zum RT anzunehmen. Neuronen der tectalen Schicht 13 projizieren exzitatorisch (glutamaterg) auf Glu R4 exprimierende Neuronen des SP. Neuronen des SP, welche das GABA synthetisierende Enzym GAD exprimieren, projizieren inhibitorisch (GABAerg) auf Neuronen des RT, welche den GABAA-Rezeptor exprimieren. Außerdem sind GABAerge Interaktionen subpraetectaler Neuronen denkbar, da ein hoher Anteil dieser Neuronen das Calcium-bindende Protein PV exprimiert.

Diskussion
93
Untersuchungen konnten Hinweise auf die Expression von GABA-Rezeptoren im
RT von Vögeln gesammelt werden. Dietl und Mitarbeiter (1988b) demonstrierten
die Synthese von GABA-Benzodiazepin-Rezeptoren im RT von Tauben, und eine
Arbeitsgruppe um Tömböl konnte in elektronenmikroskopischen immunologischen
Untersuchungen bei Hühnchen die Präsens von symmetrischen Synapsen mit
GABAergen Vesikeln an rotundalen Neuronen demonstrieren (Tömböl et al.,
1994). Die tectalen Neuronen der Schicht 13, welche direkt zum RT projizieren,
sind weder GAD- noch GABA-positiv (Karten und Mpodozis, nicht veröffentlicht;
eigene Befunde) und scheiden somit mutmaßlich als Ursprung einer
inhibitorischen Innervation aus. Anhand der vorliegenden Daten ist hingegen
anzunehmen, daß die Projektionen des SP und IPS inhibitorisch über den
Neurotransmitter GABA auf rotundalen Neuronen wirken. Parallel zu dieser
inhibitorischen Projektion der beiden Kerne auf den RT ist die vermutlich
kolokalisierte Expression von Parvalbumin und GABA in den Neuronen des SP
und IPS ein Hinweis auf intrinsische, inhibitorische Signalwege innerhalb des
SP/IPS-Komplexes. Diese Theorie wird unterstützt durch die zusätzliche
Expression von GABAA-Rezeptor im SP/IPS-Komplex. Für die Signalverarbeitung
visueller Informationen entscheidend ist aber die parallele Projektion ausgehend
vom TO zum RT. Hierbei ist der „direkte“ Weg, ausgehend von tectalen Schicht 13
Neuronen vermutlich exzitatorisch (Ngo et al., 1994, Theiss et al., 1998).
Hingegen ist der „indirekte“ Informationsfluß ausgehend von tectalen Schicht 10
und 13 Neuronen zu den praetectalen Neuronen exzitatorisch, und dann weiter zu
rotundalen Neuronen inhibitorisch, und infolge seiner polysynaptischen
Verbindungen langsamer. Außerdem ist die tecto-rotundale Projektion bilateral
(Revzin & Karten, 1967; Hunt & Künzle, 1976a), während Efferenzen des SP/IPS-
Komplexes ipsilateralen Einfluß auf den RT nehmen (Mpodozis et al., 1996;
eigene Befunde). Für die Signalverarbeitung in diesen Kerngebieten bedeutet
dies, daß tectale Neurone den RT bilateral exzitatorisch aktivieren, während der
inhibitorische Einfluß des SP/IPS-Komplexes auf den ipsilateralen RT beschränkt
bleibt.

Diskussion
94
8 Mögliche Mechanismen einer parallelen
Informationsverarbeitung
In diesem System stellt sich nun die Frage, warum visuelle Informationen auf zwei
parallelen Wegen zum RT gelangen. Bei der Betrachtung der Funktionen des RT
wird deutlich, daß es sich um einen sehr heterogen aufgebauten Kern handelt, in
welchem verschiedene visuelle Qualitäten wie Helligkeit, Farbe, Bewegungen von
Objekten in distinkten Bereichen dieses Kernes verarbeitet werden (siehe
Gliederungspunkt 7). Die skizzierten Projektionen vom TO beinhalten eine direkte,
schnelle exzitatorische, und eine indirekte, langsamere inhibitorische
Informationsleitung. Der Grund für diesen Schaltkreis könnte eine
Feinabstimmung der visuellen Information im tecto-rotundalen System sein, bei
der ein Signal zunächst schnell vom TO zum RT geleitet wird, um dann zeitlich
verzögert von Neuronen des SP/IPS-Komplexes beendet oder moduliert zu
werden. Auf das tectofugale System bezogen könnte die Rolle des SP/IPS-
Komplexes in der zeitlichen Modulation des tectofugalen Informationsflusses
liegen.
Gleichwohl sind elektrophysiologische Untersuchungen, zum Beispiel vor und
nach Läsion der praetectalen Kerne notwendig, um die vermutlich sehr
differenzierten Einflüsse (abhängig von der Lokalisation der Synapsen: axo-
dendritisch / axo-somatisch) des SP/IPS-Komplexes bei der Informations-
verabeitung im tecto-rotundalen System zu erfassen.
Auch bei Mammaliern ist eine vergleichbare Projektion in den homologen
Kerngebieten (TO = superiorer Colliculus; RT = Pulvinar) anzunehmen (Shimizu &
Karten, 1993). Zum Beispiel wurde bei Eichhörnchen gezeigt, daß der superiore
Colliculus bilateral die caudale Pulvinar innerviert, und darüber hinaus GAD-
immunreaktive Axone lokalisiert sind (Raczkowski & Diamond, 1980). Obwohl die
Quelle dieser Axone noch nicht erfaßt ist, zeigen sich deutliche Parallelen zu dem
beschriebenen System bei Vögeln, wobei auch hier Modifikationen „direkter
Projektionen“ durch „indirekte Projektionen“ vorstellbar sind.
Die in dem vorliegenden Modell dargestellten Projektionen tectaler Projektion
unter Einbeziehung der verfügbaren biochemischen Informationen deuten sehr
komplexe Mechanismen der Informationsverarbeitung im tectofugalen Sehsystem

Diskussion
95
und den entsprechend verbundenen Kerngebieten bei Vögeln an. Hierbei
erscheint es wahrscheinlich, daß eine primäre Information modulierend durch
weitere, parallele Kerngebiete beeinflußt wird. Der Grund einer solchen
Modulation liegt möglicherweise in einer schärferen Trennung verschiedener
Signale, die als Fluß an Einzelinformationen in den Kerngebieten eintreffen. Über
die Bedeutung der unterschiedlichen rostralen und caudalen Innervationen des RT
durch den SP/IPS-Komplexes kann gegenwärtig nur spekuliert werden. Auffallend
ist, daß der gesamte rostrale RT vom SP Efferenzen erhält. In diesem rotundalen
Bereich werden Informationsqualitäten wie Größe, Richtung und Geschwindigkeit
eines Objektes verarbeitet (Revzin, 1979), ebenso aber auch Informationen über
die Farbe und Helligkeit (Wang et al., 1993). Bei einer solchen Fülle
unterschiedlicher visueller Signale erscheint eine scharfe Trennung der einzelnen
Informationen notwendig. Auch im caudalen RT, der nicht komplett von Neuronen
des SP innerviert wird, sind feine Abstimmungen der eingehenden Informationen
durch modulierende Einflüsse der praetectalen Kerne sinnvoll; in diesen
rotundalen Bereichen werden Meldungen über Bewegungen von Objekten
verarbeitet (Wang et al., 1993), Informationen, die vermutlich einer ständigen
Veränderung unterliegen und sehr präzise abgestimmt werden müssen.
Die vorliegende Arbeit liefert morphologische und biochemische
Basisinformationen, um in weiteren, zum Beispiel elektrophysiologischen und
verhaltensbiologischen Untersuchungen die Funktionen der modulierenden
Einflüsse des SP/IPS-Komplexes auf den RT an Tauben zu erfassen.

Diskussion
96
9 Zusammenfassung
Im Vogelgehirn erfolgt die Verarbeitung visueller Informationen hauptsächlich über
zwei aufsteigende Sehbahnen, dem tectofugalen und dem thalamofugalen
System. Das tectofugale System verläuft von der Retina über das kontralaterale
Tectum opticum zum thalamischen Nucleus rotundus und von dort ausgehend
zum telencephalen Ectostriatum. Während die Projektionen vom Tectum opticum
zum Nucleus rotundus bereits in zahlreichen Untersuchungen beschrieben
wurden, waren mögliche Assoziationen des Nucleus subpraetectalis zum
tectofugalen System bislang weitesgehend spekulativ.
Das Ziel der vorliegenden Arbeit war es daher, die Konnektivitäten des Nucleus
subpraetectalis insbesondere zum tectofugalen System bei der Taube Columba
livia zu untersuchen. Desweiteren sollten immunhistochemische
Charakterisierungen dieses Kernes mögliche exzitatorische und/oder
inhibitorische Signalflüsse in diesem System näher analysieren.
Die Konnektivitäten wurden mit Hilfe der beiden neuronalen Tracer Biotin-Dextran-
Amin (BDA) oder der Choleratoxin ß-Untereinheit (CtB) untersucht. Die Tracer
wurden dazu in insgesamt 10 Tieren mit Hilfe einer stereotaktischen Einrichtung in
den Nucleus subpraetectalis injiziert. 3-7 Tage nach den BDA-Injektionen wurden
vornehmlich anterograde Markierungen im Nucleus rotundus detektiert, jedoch
zeigten sich auch Anfärbungen im Tectum opticum. Mit dem Tracer CtB wurden
neben den anterograden Signalen im Nucleus rotundus auch deutliche retrograde
Markierungen im Tectum opticum nach 3-5 Tagen offenbar. Am Injektionsort
wurden mit beiden Substanzen somatische Markierungen der Neuronen des
Nucleus subpraetectalis sowie des Neuropil beobachtet. Im Nucleus interstitio-
praetecto-subpraetectalis fanden sich zahlreiche Fasermarkierungen jedoch nur
vereinzelt somatische Anfärbungen. Retrograde Markierungen im Tectum opticum
waren ebenfalls durch somatische und Neuropil-Anfärbungen gekennzeichnet,
während anterograd im Nucleus rotundus vermutlich axonale Terminalien
dargestellt wurden. Außerdem konnten Konnektivitäten zum Nucleus praetectalis
aufgezeigt werden.
Bei diesen insgesamt ausschließlich ipsilateralen Markierungen wurde im Tectum
opticum nach BDA-Injektionen ein dorso-ventral zunehmender Gradient markierter

Diskussion
97
Somata in der Schicht 13 beobachtet, ohne Unterschiede in der rostro-caudalen
Ausdehnung. Im Nucleus rotundus waren hingegen caudal Terminalien im
ausschließlich lateralen Anteil dieses Kerns markiert, während im rostralen
Nucleus rotundus Terminalien im gesamten Kerngebiet beobachtet wurden.
Die immunhistochemischen Untersuchungen des Nucleus subpraetectalis dienten
desweiteren dazu, mögliche inhibitorische und exzitatorische Signalflüsse in
diesen Kernen zu analysieren. In der vorliegenden Untersuchung wurden dazu
spezifische Antikörper gegen die ? -Untereinheit des GABAA-Rezeptors, das für die
GABA-Synthese erforderliche Enzym Glutamat-Decarboxylase (GAD), die
Glutamat-Rezeptoruntereinheit GluR4, sowie das Calcium-bindende Protein
Parvalbumin eingesetzt. Somatische Markierungen von Neuronen des Nucleus
subpraetectalis wurden gezeigt für den GABAA-Rezeptor, GAD, GluR4 und
Parvalbumin, wobei die Antikörper gegen den GABAA-Rezeptor, GAD und
Parvalbumin auch das Neuropil im Nucleus subpraetectalis anfärbten.
Eine weiterhin durchgeführte quantitative Auswertung dieser Immunmarkierungen
stellte die Verteilung dieser Proteine im Bezug zur Anzahl an Neuronen in der
rostro-caudalen Ausdehnung des Nucleus subpraetectalis dar. Hierbei zeigte sich,
daß die Antikörper gegen den GABAA-Rezeptor, GAD und Parvalbumin eine
konstante Verteilung in der rostro-caudalen Ausdehnung des Nucleus
subpraetectalis aufwiesen, wobei etwa zwei-drittel aller Neurone immunpositiv auf
alle genannten Antikörper reagierten. Dagegen fand sich beim Antikörper gegen
GluR4 ein Ansteigen der immunpositiven Zellen von caudal nach rostral um den
Faktor 2,5. Es wurden keine links-rechts Unterschiede in der Expression dieser
Antigene beobachtet.
Diese Daten der Konnektivitätsuntersuchungen sowie der immunhistochemischen
Studien wurden im Bezug bereits bekannter Informationen in einem Modell zum
Einfluß des Nucleus subpraetectalis auf die mögliche Signalverarbeitung im
tectofugalen System zusammengefaßt. Demnach erhält der Nucleus
subpraetectalis direkten Eingang von tectalen Schicht 13 Neuronen. Da die
tectalen Schicht 13 Neuronen kein GAD exprimieren, während der exzitatorische
Neurotransmitter Glutamat in dieser tectalen Schicht nachgewiesen wurde, ist es
wahrscheinlich, daß der afferente Eingang des Nucleus subpraetectalis

Diskussion
98
exzitatorisch auf den GluR4 exprimierenden Neuronen terminiert. Aufgrund der
GAD und GABAA-Rezeptor Expression im Nucleus subpraetectalis sind
intrinsische inhibitorische Wechselwirkungen möglich, gleichwohl die GAD-
Synthese auch im Zusammenhang mit einer vermutlich inhibitorischen Projektion
des Nucleus subpraetectalis auf den Nucleus rotundus zu sehen ist. In der
vorliegenden Arbeit wie auch in zuvor durchgeführten Untersuchungen wurde
gezeigt, daß der Nucleus rotundus Glutamat-Rezeptoren ebenso wie GABA-
Rezeptoren synthetisiert.
Anhand dieser Daten sind zwei parallele Projektionen visueller Informationen vom
Tectum opticum zum Nucleus rotundus denkbar. Die erste, bereits ausführlich
beschriebene Projektion, verläuft direkt von den tectalen Schicht 13 Neuronen, um
exzitatorisch auf rotundalen Neuronen zu terminieren. Dieser monosynaptische
Weg ist vermutlich sehr schnell. Bei der zweiten Projektion hingegen wird
möglicherweise eine Kopie der Informationen von den tectalen Schicht 13
Neuronen zum Nucleus subpraetectalis gesandt, und anschließend in diesem über
noch nicht näher identifizierbare intrinsische Signalwege verarbeitet. Von diesem
Kerngebiet aus wird die Information anschließend zum Nucleus rotundus
weitergeleitet, vermutlich mit inhibitorischer Wirkung. Es handelt sich bei diesem
Signalweg um einen polysynaptischen Weg, der vermutlich gekennzeichnet ist
durch eine langsamere, und damit zeitlich spätere inhibitorische Wirkung auf den
Neuronen des Nucleus rotundus. Dieser Signalfluß könnte somit eine
modulatorische Wirkung des Nucleus subpraetectalis auf die Signalverarbeitung
im tectofugalen System darstellen.

Literaturverzeichnis
99
10 Literaturverzeichnis
1. Acheson, D. W., Kemplay, S. K., Webster, K. E. (1980). Quantitative analysis
of optic terminal profile distribution within the pigeon optic tectum. Journal of
Neuroscience, 5, 1067-1084.
2. Angaut, P. & Repérant, J. (1978). A light and electron microscopic study of the
nucleus isthmo-optic in the pigeon. Archiv der Anatomischen Mikroskopie, 67,
63-78.
3. Angelucci, A., Clasca, F., Sur, M. (1996). Anterograde axonal tracing with the
subunit B of cholera toxin: a highly sensitive immunohistochemical protocol for
revealing fine axonal morphology in adult and neonatal brains. Journal of
Neuroscience Methods, 65, 101-112.
4. Beart, P.M. (1976). An evaculation of L-Glutamate as the transmitter released
from optic neve terminals of the pigeon. Brain Research, 110, 99 104.
5. Benowitz, L. I. & Karten, H. J. (1976). Organization of tectofugal visual pathway
in pigeon: retrograde transport study. Journal of Comparative Neurology, 167,
503-520.
6. Binggeli, R. L. & Paule, W. J. (1969). The pigeon retina: Quantitative aspects of
the optic nerve and ganglion cell layer. Journal of Comparative Neurology, 137,
1-18.
7. Bischof, H. J. & Niemann, J. (1990). Contralateral projection of the optic tectum
in the zebra finch (Taenopygia guttata castanotis).Cell and Tissue Research,
262, 307-313.
8. Bloch, S. & Martinoya, C. (1982). Comparing frontal and lateral viewing in the
pigeon. Tachistoscopic visual acuity as a function of distance. Behavioural
Brain Research, 5, 231-244.
9. Bloch, S. & Maturana, H. R. (1971). Oil droplet distribution and colour in the
pigeon. Nature of New Biology, 234, 284-285.
10. Blough, P. M. (1971). The visual acuity of the pigeon for distant targets. Journal
of the Experimental Analysis of Behavior, 15, 57-67.
11. Bonin, B. von. (1962). Anatomical asymmetries of the cerebral hemispheres.
In: V.B.Mountcastle (Hrsg.) Interhemispheric Relations and Cerebral
dominance. Baltimore: Johns Hopkons Press.

Literaturverzeichnis
100
12. Bormann, J. (1988). Electrophysiology of GABAA and GABAB receptor
subtypes. Trends in Neuroscience, 11(3), 112-116.
13. Brandt, H. M. & Apkarian, A. V. (1992). Biotin-dextran: a sensitive anterograde
tracer for neuroanatomic studies in rat and monkey. Journal of Neuroscience
Methods, 45, 35-40.
14. Braun, K., Scheich, H., Schachner, M. & Heizmann, C.-W. (1985). Distribution
of parvalbumin, cytochromoxidase activity and 14C-2-desoxyglucose uptake in
the brain of the zebra finch. II. Visual system. Cell Tissue Research, 240, 117-
127.
15. Braun, K., Scheich, H., Zuschratter, W., Heizmann, C. W., Matute, C. & Streit,
P. (1988). Postnatal development of parvalbumin-, calbindin- and adult GABA
immunoreactivity in two visual nuclei of zebra finch. Brain Research, 475, 205-
217.
16. Cajal, S. R. Y (1911). Histologie du Système Neveux de l`Homme et des
Vertébrés, Vol. 2, Maloine, Paris.
17. Celio, M. R. & Heizmann, C. W. (1981). Calcium-binding protein parvalbumin
as a neuronal marker. Nature (London), 293, 300-302.
18. Celio, M. R. (1986). Parvalbumin in most ? -aminobutyric acid-containing
neurons of the rat cerebral cortex. Science, 231, 995-997.
19. Celio, M. R., Baier, W., Schärer, L. Gregersen, H. J., de Viragh, P. A. &
Norman, A. W. (1990). Monoclonal antibodies directed against the calcium
binding protein calbindin D-28k. Cell Calcium, 11, 599-602.
20. Chan, S. Y., Ochs, S. & Worth, R. M. (1980). The requirement for calcium ions
and the effect of other ions on axoplasmic transport in mammalian nerve.
Journal of Physiology (London), 301, 477-504.
21. Clarke, P. G. H. & Whitteridge, D. (1976). The Projection of the retina, including
the “red area“ on to the optic tectum of the pigeon. Quarterly Journal of
Experimental Physiologie, 61, 351-358.
22. Cowan, W. M., Adamson, L. & Powell, T. P. S. (1961). An experimental study
of the avian visual system. Journal of Anatomy, 95, 545-563. Crossland, W. J.,
Cowan, W. M., Rogers, L. A. & Kelly, J. P. (1974). The specification of the
retino-tectal projection in the chick. Journal of Comparative Neurology,155,
127-164.

Literaturverzeichnis
101
23. Curcio, C. A. & Allen, K. A. (1990). Topography of ganglion cells in human
retina. Journal of Comparative Neurology, 300, 5-25.
24. Deng, C. & Rogers, L.J. (1998). Organisation of the Tectorotundal and SP/IPS-
Rotundal Projections in the Chick. Journal of Comparative Neurology, 394,
171-185.
25. Dietl, M. M., Cortés, R. & Palacios, J. M. (1988a). Neurotransmitter receptors in
the avian brain. II. Muscarininc receptors. Brain Research, 439, 360-365.
26. Dietl, M. M., Cortes, R. & Palacios, J. M. (1988b). Neurotransmitter receptors in
the avian brain. III. GABA-benzodiazepine receptors. Brain Research, 439,
366-371.
27. Domenici, L., Waldvogel, H. J., Matute, C. & Streit, P. (1988). Distribution of
GABA-like immunoreactivity in the pigeon brain. Neuroscience, 25, 931-950.
28. Dunlap, K. (1981). Two types of gamma-aminobutyric acid receptor on
embryonic sensory neurones. British Journal of Pharmacology, 74(3), 579-585.
29. Eccles, J. C. & McGeer, P. L. (1979). Ionotrophic and neurotransmission.
Trends in Neuroscience, 2, 39-40.
30. Emmerton, J. (1983). Vision. In: Abs, M. (Hrsg.): Physiology and Behavior of
pigeon, Academic Press, New York, Kap. 13: 245-266.
31. Engelage, J., Bischof, H. J. (1989). Flash evoked potentials in the ectostriatum
of the zebra finch: a current source-density analysis. Experimental Brain
Research, 74, 563-572.
32. Fagg, G. E. (1985). L-Glutamate, excitatory amino acid receptors and brain
function. Trends in Neuroscience, 8, 207-210.
33. Freund, T. F. (1989). GABAergic septohippocampal neurons containing
parvalbumin. Brain Research, 478, 375-381.
34. Galifret, Y. (1968). Les diverses aires fonctionelles de la retine di pigeon.
Zeitschrift für Zellforschung und Mikroskopische Anatomie, 86, 535-545.
35. Gamlin, P. D., Reiner, A., Keyser, K. T., Brecha, N. & Karten H. J. (1996).
Projection of the nucleus pretectalis to a retinorecipient tectal layer in the
pigeon (Columba livia). Journal of Comparative Neuroscience, 368, 424-438.
36. Glick, S.D., D.A. Ross, L.B. Hough. (1982). Lateral asymmetry of
neurotransmitters in human brain. Brain Research 234; 53-63.

Literaturverzeichnis
102
37. Granda, A. M. & Yazulla, S. (1971). The spectral sensitivity of single units in
the nucleus rotundus of pigeon, Columba livia. Journal of General Physiology,
57, 363-384.
38. Granda, R. H. & Crossland, W. J. (1989). GABA-like immunoreactivity of
neurons in the chicken diencephalon and mesencephalon. Journal of
Comparative Neurology, 287(4), 455-469.
39. Greengard, P., Snyder, G., Fisone, G., Aperia, A. (1996). Interactions of signal
transduction pathways in the nervous system. Challanges and Perspectives in
Neuroscience (D. Ottoson, ed.), pp. 3-26. Pergamon, London.
40. Güntürkün, O. & Karten, H. J. (1991). An immunocytochemical analysis of the
lateral geniculate complex in the pigeon (Columba livia). Journal of
Comparative Neurology, 314(4),721-749.
41. Güntürkün, O. & Melsbach, G. (1992). Asymmetric tecto-rotundal projections in
pigeons: Cues to the origin of lateralized binocular integration. In: Elsner, N. &
D. W. Richter (Hrsg.), Rhythmogenesis in Neurons and Networks, Georg
Thieme Verlag, 1992, pp. 364.
42. Güntürkün, O., Melsbach, G. Hörster, W. & Daniel, S. (1993). Different sets of
afferents are demonstrated by the two fluorescent tracers Fast Blue and
Rhodamine. Journal of Neuroscience Methods, 49, 103-111.
43. Hahmann, U. & Güntürkün, O. (1993). The visual acuity for the lateral visual
field in the pigeon (Columba livia). Vision Research, 33, 1659-1664.
44. Hardy, O., Leresche, N. & Jassik-Gerschenfeld, D. (1985). Morphology and
laminar dirstribution of electrophysiologically identified cells in the pigeon´s
optic tectum: an intracellular study. Journal of Comparative Neurology, 233,
390-404.
45. Hayes, B. P. & Holden, A. L. (1983). The distribution of displaced ganglion cells
in the retina of the pigeon. Experimental Brain Research, 49, 181-188.
46. Hayes, B. P. & Webster, K. E. (1975). An electrone microscope study of the
retino-receptive layers of the pigeon optic tectum. Journal of Comparative
Neurology, 162, 447-466.
47. Hayes, B. P. & Webster, K. E. (1985). Cytoarchitectural fields and retinal
termination: an axonal transport study of laminar organization in the avian optic
tectum. Neuroscience, 16, 641-657.

Literaturverzeichnis
103
48. Hedblom, E., Kirkness, E.F. (1997). A novel class of GABAA receptor subunit
in tissues of the reproductive system. Journal of Biological Chemistry 272,
15346-15350.
49. Heizmann, C. W. (1984). Parvalbumin, an intrazellular calcium-binding protein;
distribution, properties and possible roles in mammalian cells, Experimentia,
40, 910-921.
50. Hellmann, B. & Güntürkün, O. (1999). Visual-field-specific heterogeneity within
the tecto-rotundal projection of the pigeon. European Journal of Neuroscience,
11, 2635-2650.
51. Hellmann, B. & Güntürkün, O. (2001). Structural organization of parallel
information processing within the tectofugal visual system of the pigeon.
Journal of Comparative Neurology, 429, 94-112.
52. Henke, H., Schenker, T. M. & Cuénod, M. (1976a). Uptake of neurotransmitter
candidates by pigeon optic tectum. Journal of Neurochemistry, 26, 125-130.
53. Henke, H., Schenker, T. M. & Cuénod, M. (1976b). Effects of retinal ablation on
uptake of glutamate, glycine, GABA, proline and choline in pigeon tectum.
Journal of Neurochemistry, 26, 131-142.
54. Heschl, R.L. (1878). Über die vordere quere Schläfenwindung des
Menschlichen Grosshirns. Wien: Braumüller.
55. Hill, D. R. & Bowery, N. G. (1981). 3H-baclofen and 3H-GABA bind to
bicuculline-insensitive GABA B sites in rat brain. Nature, 290(5802), 149-152.
56. Hirokawa, N. (1998). Kinesin and dynein superfamily proteins and the
mechanism of organelle transport. Science. 279, 519-526. Review.
57. Hodos, W. & Bonbright, J. C. Jr (1975). Intensity and pattern discrimination
after lesions of the pretectal complex, accessory optic nucleus and ventral
geniculate in pigeons. Journal of Comparative Neurology, 161, 3-18.
58. Hodos, W. & Karten, H. J. (1966). Brightness and pattern discrimination deficits
in the pigeon after lesions of nucleus rotundus. Experimental Brain Research,
2, 151-167.
59. Huang, L. H., Li, J. L. & Wang, S. R. (1998). Glutamatergic neurotransmission
from the optic tectum to the contralateral nucleus rotundus in pigeons. Brain,
Behavior and Evolution, 52, 55-60.

Literaturverzeichnis
104
60. Hunt, S. P. & Künzle, H. (1976a). Observations on the projections and
organization of the pigeon optic tectum: an autoradiographic study based
anterograde and retrograde, axonal and dendritic flow. Journal of Comparative
Neurology, 170,153-172.
61. Hunt, S. P. & Künzle, H. (1976b). Selective uptake and transport of label within
three identified neuronal systems after injection of [3H]-GABA into the pigeon
optic tectum: an autoradiographic and golgi study. Journal of Comparative
Neurology, 170, 173-190.
62. Kaufmann, D. L., McGinnis, J. F., Krieger, N. R. & tobin, A. J. (1986). Brain
glutamate Decarboxylase cloned in lambda gt-11: fusion protein produces
gamma-aminobutyric acid. Science, 232 (4754), 1138-1140.
63. Karten, H. J. & Hodos, W. (1967). A stereotaxic atlas of the brain of the pigeon
(Columba livia). The John Hopkins Press, Baltimore, Maryland.
64. Karten, H. J. & Hodos, W. (1970). Telencephalic projections of the nucleus
rotundus in the pigeon (Columba livia). Journal of Comparative Neurology, 140,
35-52.
65. Karten, H. J. & Revzin, A. M. (1966). The afferent connections of the nucleus
rotundus in the pigeon. Brain Research, 2, 368-377.
66. Karten, H. J. & Shimizu, T. (1989). The origins of neocortex: Connections and
laminations as distinct events in evolution. Journal of Cognitive Neuroscience,
1, 291-301.
67. Karten, H.J., Cox, K., Mpodozis, J. (1997). Two distinct populations of tectal
neurons have unique connections within the retinotectorotundal pathway of the
pigeon. Journal of the comparative Neurology 387, 449-465.
68. Keinänen, K., Wisden, W. & Sommer, B. (1990). A family of AMPA-selective
glutamate receptors. Science, 249, 556-560.
69. Kondo, T. (1933). Über die Verbindungen des Sehhügels mit dem
Streifenhügel beim Huhn. Aus dem Anatomischen Institut der Medizinischen
Fakultät in Okayama, 45, 797-817.
70. Krnjevic, K. (1974). Chemical nature of synaptic transmission in vertebrates.
Physiological Reviews, 54, 418-540.
71. LeMay, M. (1977). Asymmetires of the skull and handedness. Journal of the
Neurological Sciences 32; 243-253.

Literaturverzeichnis
105
72. Luksch, H., Cox, K., Karten, H.J. (1998). Bottlebrush dendritic endings and
large dendritic fields: motion-detecting neurons in the tectofugal pathway.
Journal of the Comparative Neurology 396, 399-414.
73. Mariani, A. P. & Leure-du Pree, A. E. (1978). Photoreceptors and oildroplet
colors in the red area of the pigeon retina. Journal of Comparative Neurology,
182, 821-838.
74. Martinez-de-la-Torre, M., Martinez, S. & Puelles, L. (1990).
Acetylcholinesterase-histochemical differential staining of subdivisions within
the nucleus rotundus in the chick. Anatomy and Embryology, 181, 129-135.
75. Marturana, H. R. (1962). Functional organization of the pigeon retina. 22nd
International Congreß of Physiology Science, Leyden, 170-178.
76. Marturana, H. R. & Frenk, S. (1963). Directional movement and horizontal
edge detectors in the pigeon retina. Science, 142, 977.
77. Maxwell, J. H. & Granda, A. M. (1979). Receptive fields of movement-sensitive
cells in the pigeon thalamus. In: A. M. Granda & J. H. Maxwell (Hrsg.), Neural
Mechanisms of Behavior in the Pigeon, Plenum, New York, 177-197.
78. Mayer, M. L. & Westbrook, G. L. (1987b). Permeation and block of N-methyl-D-
aspartic acid receptor channels by divalent cations in mouse cultured central
neurones. Journal of Physiology (London), 394, 501-527.
79. McGill, J. I., Powell, T. P. S. & Cowan, W. M. (1966). The retinal representation
upon the optic tectum and isthmo-optic nucleus in the pigeon. Journal of
Anatomy, 100, 5-34.
80. Miceli, D. & Repérant, J. (1985). Telencephalic afferent projections from the
diencephalon and brainstem in the pigeon. A retrograde multiple-lable
flurorescent study. Experimental Biology, 44, 71-99.
81. Morris, V. B. (1970). Symmetry in a receptor mosaic demonstrated in the chick
from the frequency, spacing and arrangement of the types of retinal receptors.
Journal of Comparative Neurology, 140, 359-398.
82. Mpodozis, J., Cox, K., Shimizu, T., Bischof, H. J., Woodson, W. & Karten, H. J.
(1996). GABAerg inputs to the nucleus rotundus (pulvinar inferior) of thepigeon
(Columbia livia). Journal of Comparative Neurology, 374, 204-222.
83. Nalbach, H.-O., Wolf Oberhollenzer, F. & Remy, M. Exploring the Image. In: H.
P. Zeigler & H.-J. Bischof (Hrsg.), Vision, Brain, and Behavior in Birds, MIT
Press, Cambridge, 1993, 25-46.

Literaturverzeichnis
106
84. Ngo, T. D., Davies, D. C., Egedi, G. Y. & Tömböl, T. (1994). A Phaseolus lectin
anterograde tracing study of the tectorotundal projections in the domestic
chick. Journal of Anatomy, 184, 129-136.
85. Ngo,T. D., Nemeth, A. & Tömböl, T. (1992). Some Data on GABA-ergic
Innervation of Nucleus rotundus in Chicks. Journal für Hirnforschung, 33 (4-5),
335-355.
86. Nicoll, R. A. (1988). The coupling of neurotransmitter receptors to ion channels
in the brain. Science, 241(4865), 545-551.
87. Nixdorf, B. E. & Bischof, H. J. (1982). Afferent connections of the ectostriatum
and visual wulst in the zebrafinch (Taeniopygia guttata castanotis Gould) - an
HRP study. Brain Research, 248, 9-17.
88. Petralia, R. S. & Wenthold, R. J. (1992). Light and electron
immunocytochemical localization of AMPA-selective glutamate receptors in the
rat brain. Journal of Comparative Neurology, 318, 329-354
89. Raczkowski, D. & Diamond, I. T. (1980). Cortical connections of the pulvinar
nucleus in Galago. Journal of Comparative Neurology, 193, 1-40.
90. Reiner, A. & Powers, A. S. (1980). The effects of extensive forebrain lesions on
visual discriminative performance in turtles (Chrysemys picta picta). Brain
Research, 192, 327-337.
91. Reiner, A., Karten, H.J. (1982). Laminar distribution of the cells of the origin of
the descending tectofugal pathways in the pigeon (columba livia). Journal of
the Comparative Neurology 204, 165-187.
92. Remy, M. & Güntürkün, O. (1991). Retinal afferents to the tectum opticum and
the n.opticus principalis thalami in the pigeon. Journal of Comparative
Neurology, 305, 57-70.
93. Revzin, A. M. & Karten, H. J. (1966 / 67). Rostral projections of the optic
tectum and the nucleus rotundus in the pigeon. Brain Research, 3, 264-276.
94. Revzin, A. M. (1979). Functional localization in the nucleus rotundus. In: A. M.
Granda & J. H. Maxwell (Hrsg.). Neural Mechanisms of Behavior in the Pigeon.
Plenum Press, New York, 1979, 165-175.
95. Shimizu,T., Karten, H. J. & Woodson, W. (1988). GABAergic inputs to the
nucleus rotundus in pigeon (Columba livia). Society of Neuroscience
?Abstract?, 397.9.

Literaturverzeichnis
107
96. Shimizu, T. & Karten, H. J. The avian visual system and the evolution of the
neocortex. In: H. P. Zeigler & H.-J. Bischof (Hrsg.), Vision, Brain, and Behavior
in Birds, pp 104-114, The MIT Press, Cambridge, 1993.
97. Shimizu, T., Cox, K., Karten, H. J. & Britto, L. R. G. (1994). Cholera toxin
mapping of retinal projections in pigeons (Columba livia), with emphasis on
retinohypothalamic connections. Visual Neuroscience, 11, 441-446.
98. Sieghart, W. (1995). Structure and pharmacology of gamma-aminobutyric acid
A receptor subtypes. Pharmacological Review 47, 181-234.
99. Sladeczek, F., Pin, J. P., Recasens, M. Bockaert, J. & Weiss, S. (1985).
Glutamate stimulates inositol phosphate formation in striatal neurons. Nature,
317, 717-719.
100. Smith, S. & Augustine, G. J. (1988). Calcium ions, active zones and synaptic
transmitter release. Trends in Neuroscience, 11, 458-465.
101. Theiss, C., Hellmann, B. & Güntürkün, O. (1998). The differential distribution
of AMPA-receptor subunits in the tectofugal system of the pigeon. Brain
Research, 785, 114-128.
102. Tömböl, T. Ngo, T. D. & Egedy, G. (1992). EM and EM golgi study on
structure of nucleus rotundus in chicks. Journal für Hirnforschung, 33, 215-234.
103. Tömböl, T., Egedi, G. & Nemeth, A. (1994). Phaseolus vulgaris lectin labelled
and GABA immunogold stained terminals in nucleus rotundus. An EM study.
Journal für Hirnforschung, 35(2), 233-252.
104. Tömböl, T., Nemeth, A., Sebesteny, T. & Alpar, A. (1999). Electron
microscopic data on the neurons of nuclei subpretectalis and posterior-ventralis
thalami. A combined immunohistochemical study. Anatomical Embryology, 199,
169-183.
105. Uhlrich, D. J., Blough, P. M. & Blough, D. S. (1982). The pigeon´s distant
visual acuity as a function of viewing angle. Vision Research, 22, 429-431.
106. Van Gehuchten, A. (1892). La structure des lobes optiques chez l´embryon
de poulet. La Cellule, 8, 1.
107. Varela, F. J., Palacios, A. G. & Goldsmith, T. H., Color Vision of Birds, In:
Vision, Brain & Behavior in Birds, H. P. Zeigler & H.-J. Bischof (Eds.), MIT
Press, 1993, pp. 77-98.

Literaturverzeichnis
108
108. Veenman, C. L. & Reiner, A. (1994). The distribution of GABA-containing
perikarya, fibers, and terminals in the forebrain and midbrain of pigeons, with
particular reference to the basal ganglia and its projection targets. Journal of
Comparative Neurology, 339, 209-250.
109. Veenman, C. L., Albin, R. L., Richfield, E. K. & Reiner. A. (1994).
Distributions of GABAA, GABAB, and benzodiazepine receptors in the forebrain
and midbrain of pigeons. Journal of Comparative Neurology, 344, 161-189.
110. Wachtler, K. (1985). Regional distribution of muscarinic acetylcholine
receptors in the telencephalon of the pigeon (Columba livia). Journal für
Hirnforschung, 26, 85-89.
111. Wang, Y. & Frost, B. J. (1992). Time to collision is signalled by neurons in the
nucleus rotundus of pigeons. Nature, 356, 236-238.
112. Wang, Y., Jiang, S. & Frost, B. J. (1993). Visual processing in pigeon nucleus
rotundus: luminance, color, motion, and looming subdivisions. Visual
Neuroscience, 10, 21-30.
113. Wässle, H. & Boycott, B. B. (1991). Functional architecture of the mammalian
retina. Watkins, J. C. & Evans, R. H. (1981). Excitatory amino acid transmitters.
Annual Review of Pharmacology, 21, 165-204.
114. Wenthold, R. J., Hunter, C., Wada, K. & Dechesne, C. J. (1990). Antibodies
to a C-terminal peptide of the rat brain glutamate receptor subunit, GluR-A,
recognize a subpopulation of AMPA binding sites but not kainate sites. FEBS
Letter, 276, 147-150.
115. Wenthold, R. J., Yokotani, N., Doi, K. & Wada, K. (1992). Immunochemical
characterisation of the non-NMDA glutamate receptor using subunit-specific
antibodies. Journal of Biological Chemistry, 267, 501-507.
116. Whiting, P.J. (1999). The GABAA receptor gene family: new targets for
therapeutic intervention. Neurochemistry international 34, 387-390.
117. Whiting, P.J., McAllister, G., Vasilatis, D., Bonnert, T.P., Heavens, R.P.,
Smith, D.W.,Hewson, L., O´Donnell, R., Rigby, M.R., Sirinathsinghji, D.J.,
Marshall, G., Thompson, S.A., Wafford, K.A. (1997). Neuronal restricted RNA
splicing regulates the expression of a novel GABA-A receptor subunit conferring
atypical functional properties. Journal of Neuroscience 17, 5027-5037.

Literaturverzeichnis
109
118. Whiting, P.J., McKernan, R.M., Wafford, K.A. (1995). Structure and
pharmacology of vertebrate GABA-A receptor subtypes. International Rev.
Neurobiology 38, 95-138.
119. Wild, J.M. (1989). Pretectal and tectal projections to the homologue of the
dorsal lateral geniculate n. in the pigeon: an antero-retrograde tracing study with
cholera. Brain Research, 479, 130-137.
120. Young, S. R. & Martin, G. R. (1984). Optics of retinal oil droplets: a model of
light collection and polarization detection in the avian retina. Vision Research,
24, 129-137.

Danksagung Herrn Prof. Dr. O. Güntürkün möchte ich meinen Dank ausdrücken für die
Bereitstellung des Dissertationsthemas sowie für die hervorragende Betreuung
während der gesamten Anfertigung der vorliegenden Doktorarbeit.
Ebenso möchte ich meinen Dank an Herrn Prof. Dr. U. Eysel aussprechen, der als
Betreuer an der medizinischen Fakultät das Fortkommen dieser Arbeit
aufmerksam verfolgt und unterstützt hat.
Bedanken möchte ich mich besonders bei Dr. Burkhard Hellmann, der durch seine
intensive Betreuung und konstruktive Kritik eine große Unterstützung während der
gesamten Zeit bei der Fertigstellung dieser Doktorarbeit war.
Danken möchte ich auch Ariane Schwarz für ihre Hilfe in labortechnischen Fragen.
Besonders bedanken möchte ich mich bei meinen Schwiegereltern Hildegard und
Herbert Theiß, die durch ihre liebevolle Kinderbetreuung die endgültige
Fertigstellung dieser Arbeit ermöglicht haben.
Ein großer Dank gilt meinen Eltern Prof. Dr. K. Meller und C. Zaragoza Mifsud de
Meller für ihre fachliche wie familiäre Unterstützung während des gesamten
Studiums und der Zeit als Ärztin im Praktikum.
Ein besonderer Dank gilt meinem Mann Carsten.





























