Embed Size (px)
Citation preview

Das Phänomen „Leben“ ist geprägt von intensivenInteraktionen, die auf verschiedenen Komplexi-
tätsebenen stattfinden. Von besonders großer Bedeu-tung sind die Wechselwirkungen, die zwischen einemOrganismus und seiner Nahrung stattfinden. Ernäh-rung ist die Voraussetzung für Überleben, und diegrundsätzliche Überlebensfähigkeit des Einzelnen istdie logische Voraussetzung für das Überleben eines Kollektivs und somit einer ganzen Spezies.
Der Mensch stellt keine Ausnahme von diesem Prin-zip dar. Aber er nimmt eine Sonderstellung ein, dennauf Grund seiner zahlreichen kulturellen Errungen-schaften hat keine andere Spezies derart starken Ein-fluss auf ihre Nahrungsorganismen und Nahrungsbe-standteile genommen – und somit auch auf sich selbst– wie Homo sapiens.
Bezüglich des wissenschaftlichen Erkenntnisge-winns erscheint die Erforschung dieser besonderen Interaktionen zwischen den Menschen und ihrer Nah-
Der Mensch verändert die Genetik seinerNahrungsorganismen und die Nahrunggreift in die Genetik des Menschen ein. Seiteinigen Jahren wächst die Gewissheit, dassdie Komplexität dieser Wechselwirkungenwesentlich größer ist als bislang vermutet.Die jungen Forschungsdisziplinen Nutrigene-tik und Nutrigenomik tragen dabei nicht nur zur Aufklärung allgemeiner Interaktions-wege bei, sondern offenbaren zudem sowohl ethnische als auch individuelle Unterschiede.
Die Genetik der Ernährung:
Wie sich Mensch undNahrung beeinflussenFRITZ HÖFFELER
DOI:10.1002/ biuz.201410528
52 | Biol. Unserer Zeit | 1/2014 (44) © 2014 Wiley-VCH Verlag GmbH & Co. KGaA, WeinheimOnline-Ausgabe unter:wileyonlinelibrary.com
A B B . 1 Haupt-wege der Wech-selwirkungenzwischenMensch undNahrungsorga-nismus.
gezielte Veränderung der genetischen Ausstattungunserer Nahrungsorganismen(Züchtung)
Einfluss der Nahrungsbestandteile auf dieRegulation unserer Gene(Nutrigenomik)
Einfluss unserer genetischen Ausstattung auf unsere grundsätzlichen Ernährungsmöglichkeiten(Nutrigenetik)
gegenseitige Abhängigkeit zwischen bestimmtenkulturellen Neuerungen und speziellen Mutationenunserer Gene (Coevolution)

G E N E T I K U N D E R N Ä H R U N G | I M FO KU S
rung als besonders fruchtbar. Undda die Nahrung und Ernährung imZentrum zahlreicher sozialer, politi-scher und medizinischer Problemeder heutigen Menschheit stehen,könnten sich die gewonnenen Er-kenntnisse als sehr hilfreich für ver-schiedene Lösungsansätze erwei-sen.
Die Generierung und Interpre-tation riesiger Datenmengen führtezur Identifizierung von vier ver-schiedenen Wechselwirkungen: Nu -trigenetik, Coevolution, Nutrigeno-mik und Züchtung (Abbildung 1).
Nutrigenetik: der Einfluss unserer Gene
Nutrigenetik ist eine recht jungeDisziplin, in der untersucht wird,wie unsere individuelle Gen-Aus-stattung auf die zugeführte Nah-rung reagiert; ein besonderes Au-genmerk ist dabei auf die in der Bevölkerung vorkommenden Poly-morphismen gerichtet. In der Nutri-genetik wird versucht, den Zusammenhang zwischendiesen Variationen und den unterschiedlichen Reaktio-nen auf Nährstoffe zu ergründen, besonders auch imHinblick auf die Ursachen von Unverträglichkeiten undauf die Entstehung verschiedener Erkrankungen.
Beispiel 1: Abbau von AlkoholEthanol wird hauptsächlich in den Leberzellen abge-baut und zwar durch zwei Enzyme. Die Alkohol-De-hydrogenase (ADH) wandelt Ethanol reversibel inAcetaldehyd um, und die Aldehyd-Dehydrogenase(ALDH) oxidiert Acetaldehyd weiter zu Acetat, das in Acetyl-CoA eingebaut wird und für zahlreicheStoffwechselprozesse verwendet werden kann (Ab-bildung 2).
ADH kommt sowohl als Homo- als auch als Hetero-dimer vor. Für die Untereinheiten besitzt der Menschzahlreiche Genvarianten, die in fünf Klassen eingeteiltwerden (Abbildung 3). Die Klasse I nimmt eine Son-derstellung ein, denn das ADH1-Gen kommt in drei Va-riationen (A, B und C) vor. In den ADH1B- und ADH1C-Genen wurden einige Mutationen identifiziert, durchdie wir uns voneinander unterscheiden: Das GenADH1B gibt es in drei und ADH1C in zwei Varianten.Diese als Allozyme bezeichneten Modifikationen wer-den durch angehängte Ziffern voneinander unterschie-den.
Je nach dem, aus welchen Untereinheiten sich dieAlkohol-Dehydrogenasen zusammensetzen, unterschei-den sie sich in ihrer Leistungsfähigkeit zum Teil erheb-
© 2014 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.biuz.de 1/2014 (44) | Biol. Unserer Zeit | 53
CoAS
O
CH3C
Acetyl-CoA
CH
H
H
HOC
H
H
CH
H
H
C
O
H
Acetaldehyd(toxisches Zwischenprodukt)
Ethanol
Alkohol-Dehydrogenase
Aldehyd-Dehydrogenase
ABB. 2 A B BAU VO N E T H A N O L
Klasse I
ADH1C ADH1B ADH1A
ADH1C*1ADH1C*2
ADH1B*1ADH1B*2ADH1B*3
Klasse IVADH4
Klasse VADH5
Klasse IIADH2
Klasse IIIADH3
ABB. 3 O RG A N I SAT I O N D E R ADH- G E N E
TA B . K I N E T I K U N D V E R B R E I T U N G E I N I G E R ADH- G E N E
Gen-Variante Katalytische Häufigkeit (%) unterEffizienz Ost-Asiaten Kaukasiern afrikanischen amerikanischen taiwanesischer
Amerikanern Indianern Urbevölkerung
ADH1A 0,079ADH1B*1 10 ca. 30 ca. 90 84 93 9ADH1B*2 2,6 ca. 70 ca. 10 0 1 91ADH1B*3 0,056 0 0 16 6 0ADH1C*1 – ca. 90 ca. 60 85 56 99ADH1C*2 – ca. 10 ca. 40 15 44 1ADH2 0,020ADH4 0,30Die katalytische Effizienz ergibt sich aus der Umsatzgeschwindigkeit (Vmax) dividiert durch die Halbsättigung des Enzyms (Km). Die Werte wurdenan Homodimeren ermittelt.

lich voneinander (siehe Tabelle) [2]. Beispielsweise arbeitet das ADH1B1-Homodimer um das 500fache ef-fektiver als das ADH2-Homodimer. Auffällig ist die geo-graphisch/ethnische Ungleichverteilung der Allozyme.Das für die effektivste Untereinheit codierende GenADH1B*1 ist unter der nord- und westeuropäischenBevölkerungsgruppe zu über 90 % verbreitet, aber nurzu ungefähr 30 % unter der ostasiatischen Bevölkerungund zu knapp 10 % unter der taiwanesischen Urbevöl-kerung. Das Verteilungsmuster der deutlich weniger ef-fektiven Variante ADH1B2 ist nahezu umgekehrt: In dereuropäischen Bevölkerung hat dieses Gen eine Häufig-keit von circa 10 %, unter Ostasiaten von ungefähr 70 %und bei Taiwanesen von über 90 % [3].
Bei der Verarbeitung von Acetaldehyd ist haupt-sächlich ALDH2 beteiligt, ein Homotetramer. Das Genfür das Monomer trägt die Bezeichnung ALDH2*1.Auch für dieses Gen gibt eine durch Mutation entstan-dene Variante, die mit ALDH2*2 gekennzeichnet wird.Das entsprechende Protein unterscheidet sich zwarnur in einer Aminosäure vom ursprünglichen Protein,bildet jedoch, zum Tetramer zusammengesetzt, ein na-hezu inaktives Enzym. Die Konsequenzen für den Be-troffenen sind eine stark erhöhte Alkoholunverträglich-keit, da die rapide ansteigende Acetaldeyd-Konzentra-tion im Blut neben den typischen Kater-Symptomenauch Vergiftungserscheinungen auslöst, wie zum Bei-spiel das Flush-Syndrom, ein neuronal ausgelöstes,schnelles, unangenehmes Erröten der Gesichtshautund des Halses. Das Gen ALDH2*2 kommt in der euro-päischen Bevölkerungsgruppe nur sehr selten vor, istaber zu ungefähr 50 % unter Chinesen und Japanernverbreitet [3].
Die meisten Ostasiaten tragen also zwei am Alko-hol-Abbau beteiligte Enzymvarianten, die auf Grund ihrer Ineffektivität für eine hohe Alkoholsensibilität ver-antwortlich sind. Umgehen können Betroffene diesesProblem lediglich, indem sie ihren Alkoholkonsumstark einschränken.
Beispiel 2: Fructose-UnverträglichkeitFructose ist ungefähr doppelt so süß wie Glucose undwird daher verstärkt in der Lebensmittelindustrie ein-gesetzt. Mit Hilfe des Enzyms Glucose-Isomerase wirdin großtechnischem Maßstab glucosehaltiger Maissirupzu einem High-Fructose Corn Syrup (HFCS) umgewan-delt. Vor einigen tausend Jahren haben unsere Vorfah-ren mit dem Verzehr von Obst, Honig, Gemüse, Weizenoder Knollen- und Wurzelfrüchten vermutlich ungefähr20 g am Tag aufgenommen. Heute jedoch nimmt eindurchschnittlicher Amerikaner die vierfache Menge zusich.
Bei schätzungsweise bis zu einem Drittel der Bevöl-kerung westlicher Staaten stellen sich nach der Auf-nahme von nur geringen Mengen Fructose deutlicheVerdauungsbeschwerden ein, die als Fructose-Malab-
sorption bezeichnet werden. Die Ursache dafür liegtvermutlich in den Kapazitätsgrenzen des Fructose-Transporters GLUT-5, der von bestimmten Epithelzel-len des Dünndarms produziert wird [4]. Die Transport-proteine befördern Fructosemoleküle aus dem Darm-lumen in die Epithelzelle. Ein durchschnittlicherErwachsener ist in der Lage, 25 bis 50 g Fructose proMahlzeit über die Fructose-Transporter aufzunehmen;größere Mengen verursachen Verdauungsproblemewie Durchfall, Blähungen und Bauchschmerzen. Offen-sichtlich ist unser Dünndarm nicht in der Lage, beliebiggroße Mengen an GLUT-5 herzustellen und beliebiggroße Mengen Fructose zu resorbieren. Die nicht re-sorbierte Fructose verursacht im Dünn- und Dickdarmeinen osmotischen Sog und zieht Wasser aus den um-liegenden Gefäßen an (die Folge: Diarrhoe). DieFructose wird von der Bakterienflora des Dickdarmsverstoffwechselt, wobei Gase wie Wasserstoff, Kohlen-dioxid und Methan entstehen (die Folge: Blähungenund Schmerzen). Warum einige Menschen bereits nachder Aufnahme von nur 1 g Fructose Verdauungspro-bleme bekommen, andere erst nach 20 g und wiederandere offenbar gar nicht, ist bis heute noch immernicht völlig geklärt. Mutationen im GLUT5-Gen wurdennicht entdeckt, was bedeutet, dass die unterschiedli-chen Symptome nicht auf defekte Transport-Proteinezurückzuführen sind. Es wird vermutet, dass die Mengeder synthetisierten Transporter nicht bei allen Men-schen gleich ist.
Behandlungsmöglichkeiten gegen eine Fructose-Malabsorption gibt es nicht, aber eine Reduktion derzugeführten Menge kann den Beschwerdegrad erheb-lich mindern.
Coevolution: gegenseitige AbhängigkeitenPhänomene der Coevolution stellen Sonderfälle derNutrigenetik dar: Bestimmte Genvarianten haben einendeutlichen Einfluss auf spezielle Ernährungsgewohn-heiten, und umgekehrt sind eben diese Ernährungsge-wohnheiten die Ursache für die positive Selektion derGenvarianten.
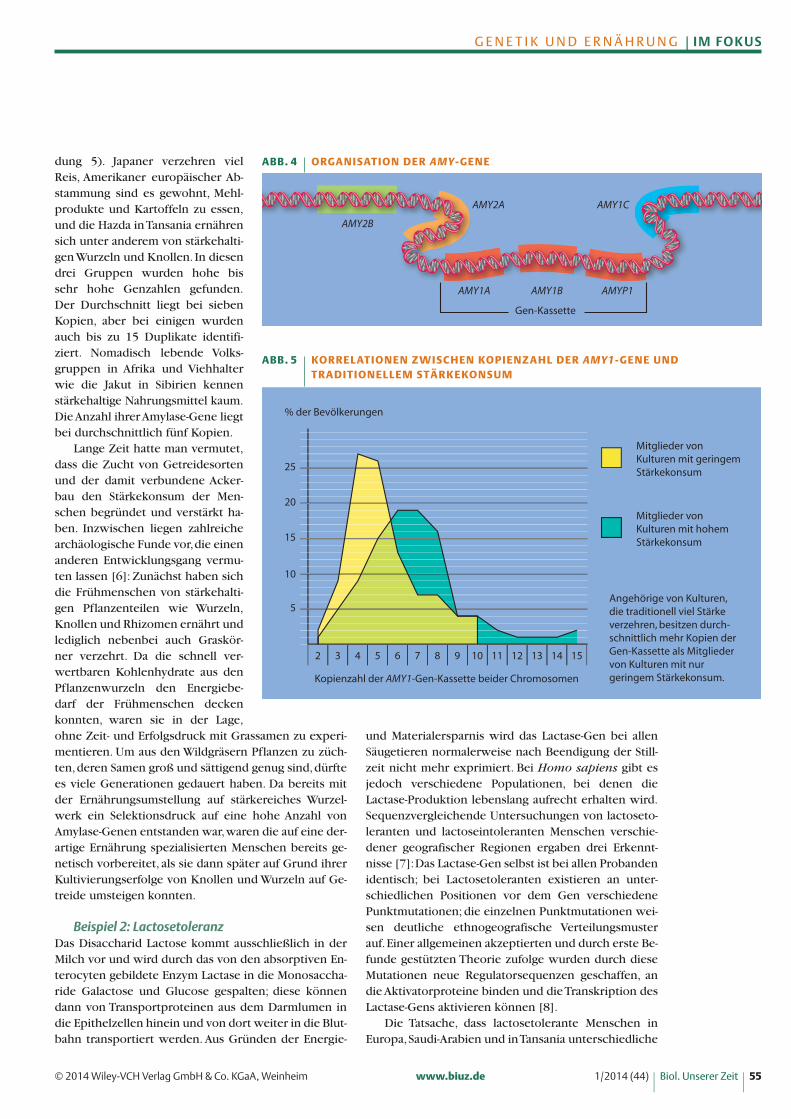
Beispiel 1: Stärkeverdauungα-Amylasen werden in der Bauch- und in den Mund-speicheldrüsen produziert. Das Enzym kommt beimMenschen in mehreren Isoformen vor. Im Pankreas wer -den die Varianten AMY2A und AMY2B transkribiert, in den Mundspeicheldrüsen die Varianten AMY1A,AMY1B und AMY1C. Die Gene AMY1A und AMY1Bbilden zusammen mit dem Pseudogen AMYP1 eineGen-Kassette, die im Verlauf der Humanevolution bis zu14-mal vervielfältigt wurde (Abbildung 4).
Die Anzahl der AMY1-Kopien unterscheidet sichbei Menschen aus unterschiedlichen Regionen derWelt sehr deutlich voneinander und sie korreliert mitder Menge des traditionellen Stärkeverzehrs [5] (Abbil-
54 | Biol. Unserer Zeit | 1/2014 (44) www.biuz.de © 2014 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
G E N E T I K U N D E R N Ä H R U N G | I M FO KU S
dung 5). Japaner verzehren vielReis, Amerikaner europäischer Ab-stammung sind es gewohnt, Mehl-produkte und Kartoffeln zu essen,und die Hazda in Tansania ernährensich unter anderem von stärkehalti-gen Wurzeln und Knollen. In diesendrei Gruppen wurden hohe bissehr hohe Genzahlen gefunden.Der Durchschnitt liegt bei siebenKopien, aber bei einigen wurdenauch bis zu 15 Duplikate identifi-ziert. Nomadisch lebende Volks-gruppen in Afrika und Viehhalterwie die Jakut in Sibirien kennenstärkehaltige Nahrungsmittel kaum.Die Anzahl ihrer Amylase-Gene liegtbei durchschnittlich fünf Kopien.
Lange Zeit hatte man vermutet,dass die Zucht von Getreidesortenund der damit verbundene Acker-bau den Stärkekonsum der Men-schen begründet und verstärkt ha-ben. Inzwischen liegen zahlreichearchäologische Funde vor, die einenanderen Entwicklungsgang vermu-ten lassen [6]: Zunächst haben sichdie Frühmenschen von stärkehalti-gen Pflanzenteilen wie Wurzeln,Knollen und Rhizomen ernährt undlediglich nebenbei auch Graskör-ner verzehrt. Da die schnell ver-wertbaren Kohlenhydrate aus denPflanzenwurzeln den Energiebe-darf der Frühmenschen deckenkonnten, waren sie in der Lage,ohne Zeit- und Erfolgsdruck mit Grassamen zu experi-mentieren. Um aus den Wildgräsern Pflanzen zu züch-ten, deren Samen groß und sättigend genug sind, dürftees viele Generationen gedauert haben. Da bereits mitder Ernährungsumstellung auf stärkereiches Wurzel-werk ein Selektionsdruck auf eine hohe Anzahl vonAmylase-Genen entstanden war, waren die auf eine der-artige Ernährung spezialisierten Menschen bereits ge-netisch vorbereitet, als sie dann später auf Grund ihrerKultivierungserfolge von Knollen und Wurzeln auf Ge-treide umsteigen konnten.
Beispiel 2: LactosetoleranzDas Disaccharid Lactose kommt ausschließlich in derMilch vor und wird durch das von den absorptiven En-terocyten gebildete Enzym Lactase in die Monosaccha-ride Galactose und Glucose gespalten; diese könnendann von Transportproteinen aus dem Darmlumen indie Epithelzellen hinein und von dort weiter in die Blut-bahn transportiert werden. Aus Gründen der Energie-
und Materialersparnis wird das Lactase-Gen bei allenSäugetieren normalerweise nach Beendigung der Still-zeit nicht mehr exprimiert. Bei Homo sapiens gibt esjedoch verschiedene Populationen, bei denen dieLactase-Produktion lebenslang aufrecht erhalten wird.Sequenzvergleichende Untersuchungen von lactoseto-leranten und lactoseintoleranten Menschen verschie-dener geografischer Regionen ergaben drei Erkennt-nisse [7]: Das Lactase-Gen selbst ist bei allen Probandenidentisch; bei Lactosetoleranten existieren an unter-schiedlichen Positionen vor dem Gen verschiedenePunktmutationen; die einzelnen Punktmutationen wei-sen deutliche ethnogeografische Verteilungsmusterauf. Einer allgemeinen akzeptierten und durch erste Be-funde gestützten Theorie zufolge wurden durch dieseMutationen neue Regulatorsequenzen geschaffen, andie Aktivatorproteine binden und die Transkription desLactase-Gens aktivieren können [8].
Die Tatsache, dass lactosetolerante Menschen inEuropa, Saudi-Arabien und in Tansania unterschiedliche
© 2014 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.biuz.de 1/2014 (44) | Biol. Unserer Zeit | 55
AMY2B
AMY2A AMY1C
AMY1A AMY1B AMYP1
Gen-Kassette
ABB. 4 O RG A N I SAT I O N D E R AMY- G E N E
2 3 4 5 6 7 8 9 10 11 12 13 14 15
5
10
15
20
25
% der Bevölkerungen
Kopienzahl der AMY1-Gen-Kassette beider Chromosomen
Angehörige von Kulturen, die traditionell viel Stärke verzehren, besitzen durch-schnittlich mehr Kopien der Gen-Kassette als Mitglieder von Kulturen mit nur geringem Stärkekonsum.
Mitglieder von Kulturen mit geringem Stärkekonsum
Mitglieder von Kulturen mit hohem Stärkekonsum
ABB. 5 KO R R E L AT I O N E N Z W I S C H E N KO PI E N Z A H L D E R AMY1- G E N E U N D T R A D I T I O N E L L E M S T Ä R K E KO N S U M

Mutationen tragen, die jeweils mit dem Auftreten derLactosetoleranz korrelieren, lässt darauf schließen, dassdiese Eigenschaft mehrfach und unabhängig voneinan-der entstanden ist. Diese Vermutung wird gestütztdurch zahlreiche archäologische Funde aus verschie-denen Regionen und Phasen der Steinzeit. Offenbarentwickelten sich die Züchtung milchproduzierenderTiere und die Herstellung von Milchprodukten inEuropa, Saudi-Arabien und Afrika unabhängig vonei-nander. Einige wenige lactosetolerante Menschen be-gannen auf gut Glück mit der Milchviehhaltung unddem Milchverzehr, der sich ernährungsphysiologischsehr positiv auswirkte – besonders auf die Entwicklungder Kinder. Es entstand innerhalb solcher Gemein-schaften ein hoher Selektionsdruck auf die mutiertenSequenzen, die sich dann innerhalb der Kollektiveschnell ausbreiteten. Als sukzessive immer mehr Er-wachsene Lactose tolerieren konnten, wurde es sinn-voll, die Milchviehhaltung zu fördern und die Milchver-arbeitung weiterzuentwickeln.
Nutrigenomik: der Einfluss unserer NahrungDie Nutrigenomik beschäftigt sich mit den Auswirkun-gen unserer Nahrungsbestandteile auf zelluläre, geneti-sche und biologische Systeme. Sie untersucht insbe-sondere die durch die Nährstoffe ausgelösten Reakti-onskaskaden unseres Stoffwechsels und stellt daherden komplementären Ansatz zur Nutrigenetik dar. Der-artige Erkenntnisse helfen festzustellen, worin die Un-terschiede zwischen einem gesunden und einem kran-ken Organismus liegen und welche Prozesse einer Er-krankung bzw. einer Genesung zugrundeliegen.
Beispiel 1: Glucose als SignalgeberGlucose ist eines der häufigsten Biomoleküle. Sie fun-giert, ebenso wie die unten beschriebenen Fette undFettsäuren, sowohl als Baumaterial als auch als Energie-lieferant. Bei nahezu allen heterotrophen Organismenentstanden im Laufe der Evolution Mechanismen, diedie Glucose selbst als Signalgeber für zahlreiche Glu-cose-verarbeitende Prozesse verwenden.
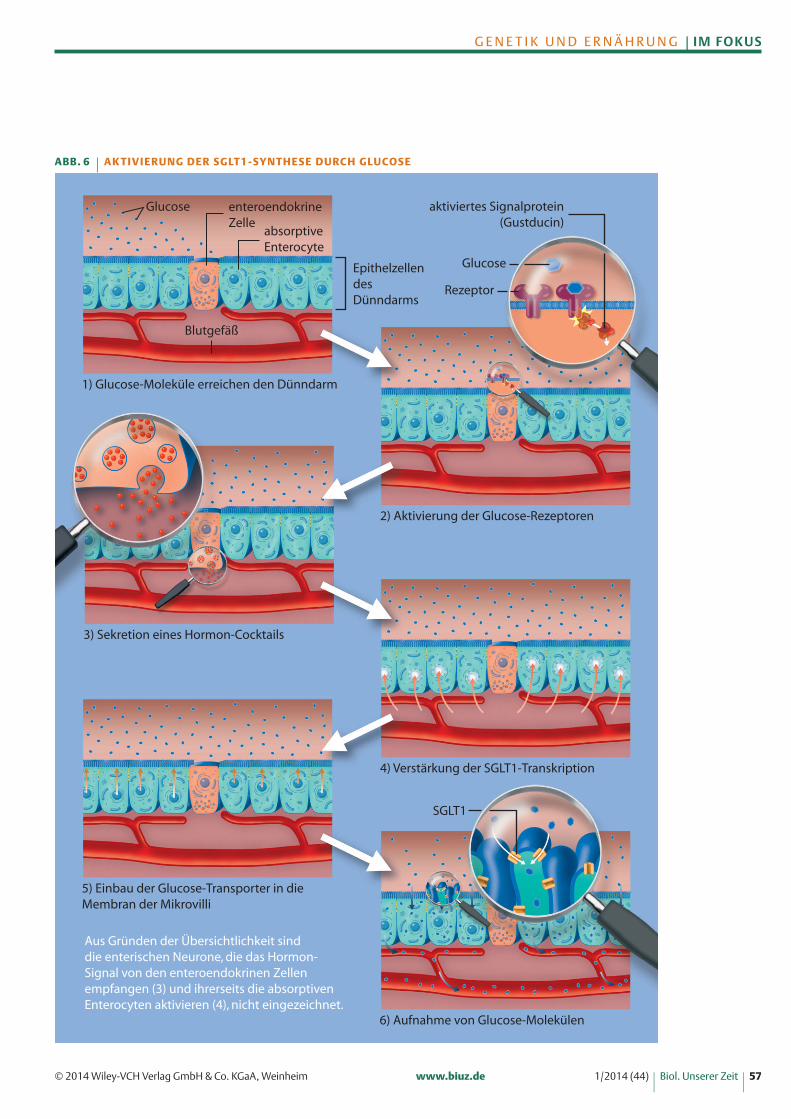
Dieselben Süßrezeptoren, die auch die Sinneszellenunserer Geschmacksknospen auf der Zunge tragen,werden von speziellen Dünndarmzellen gebildet, denenteroendokrinen Zellen. Wenn durch die Nahrung zu-geführte Glucosemoleküle an diese Rezeptoren bin-den, wird eine Reaktionskaskade in den enteroendokri-nen Zelle ausgelöst, an deren Ende ein Hormoncocktailausgeschüttet wird, der den Organismus auf die Auf-nahme und Weiterverarbeitung neuer Zuckermolekülevorbereitet (Abbildung 6) [9]. Eines der Hormone istdas Glucagonähnliche Peptid 2 (GLP-2). Gemäß einerweithin akzeptierten Theorie bindet das Hormon anRezeptoren enterischer (dem Darm zugehöriger) Ner-venzellen, die das Signal den Darm entlang weiterleitenund dann ihrerseits Signalmoleküle ausschütten. Diese
docken an Rezeptoren der absorptiven Enterocyten an,einer weiteren Gruppe von Dünndarmzellen. Die Akti-vierung setzt in den Darmzellen eine Reaktionskette inGang, durch die letztlich das Gen für den Glucose-transporter SGLT1 verstärkt transkribiert wird [10]. DieGlucosetransporter werden in die dem Darm zuge-wandte Zellmembran eingebaut, wodurch die absorpti-ven Enterocyten nun wesentlich schneller die Gluco-semoleküle aus dem Darminhalt aufnehmen und an dieBlutbahn weiterleiten. Glucose beschleunigt also ihreeigene Resorption aus dem Darm.
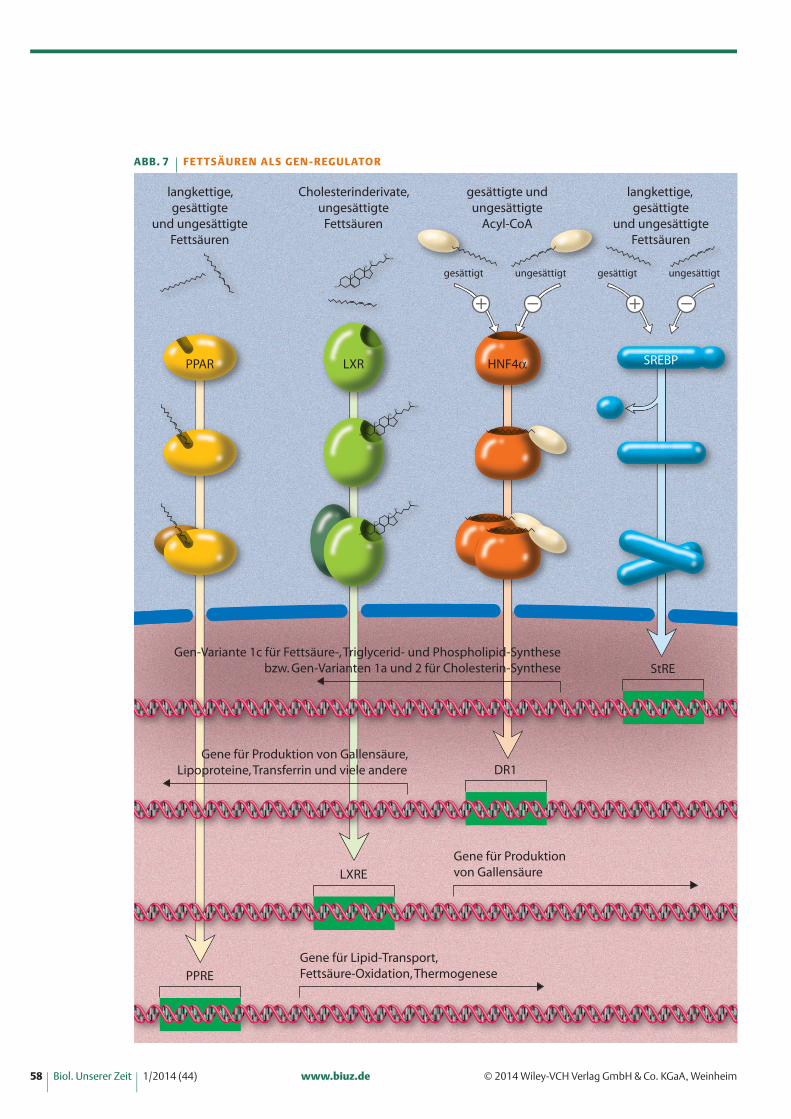
Beispiel 2: Fettsäuren als SignalgeberFettsäuren sind sowohl in der Lage, über die Aktivie-rung von Rezeptoren Hormonsignale an verschiedeneZelltypen auszusenden, als auch durch Interaktionenmit Transkriptionsfaktoren die Transkriptionsrate zahl-reicher Gene zu regulieren (Abbildung 7) [12, 13]. Sokönnen mehrfach ungesättigte Fettsäuren die Lipid-Synthese in der Leber drosseln, die Fettsäure-Oxidationin der Leber und in den Muskeln ankurbeln und dieSpeicherung von Glycogen aktivieren. Mindestens vierTranskriptionsfaktoren können durch Fettsäuren be-einflusst werden: Peroxisome Proliferator-Activated Re-ceptor (PPAR), Liver X Receptor (LXR), Hepatic Nu-clear Factor 4α (HNF-4α) und Sterol Regulatory Ele-ment Binding Protein (SREBP).
PPAR besitzt eine Bindetasche für eine Fettsäure; la-gert sich diese dort an, entsteht eine kleine Konfigura-tionsveränderung des Proteinkomplexes, die zur Folgehat, dass sich der Komplex zusammen mit einem wei-teren Protein an eine als Peroxisome Proliferator Re-sponsive Element (PPRE) bezeichnete Regulationsse-quenz bindet. Durch PPREs regulierte Gene codierenProteine, die am Lipidtransport, an der Fettsäure-Oxi-dation und an der Thermogenese (der Wärmeentwick-lung durch Fettverbrennung in der Leber und im brau-nen Fettgewebe) beteiligt sind.
LXR kann durch verschiedene Cholesterinderivateund durch langkettige Fettsäuren aktiviert werden.Auch dieser Rezeptor bildet zusammen mit einem zu-sätzlichen Faktor ein Heterodimer, das an charakteristi-sche Sequenzmotive binden kann, die als LXR Respon-sive Element (LXRE) bezeichnet werden. Die durchLXR regulierten Gene exprimieren Produkte, die dieZelle vor einer Cholesterin-Überladung schützen, unteranderem durch die Synthese und Ausschleusung vonGallensäuren.
HNF-4α ist ein Transkriptionsfaktor, der je nach Fett-säuretyp aktiviert oder gehemmt werden kann. An ihnbinden durch Acetyl-CoA aktivierte Fettsäuren (Acyl-CoAs). Gesättigte Acyl-CoAs aktivieren, mehrfach unge-sättigte dagegen hemmen den Faktor. HNF-4α kontrol-liert direkt oder indirekt die Expression verschiedenerGene, wie zum Beispiel solche für Lipoproteine, für dasam Eisenstoffwechsel beteiligte Transferrin, für die Syn-
56 | Biol. Unserer Zeit | 1/2014 (44) www.biuz.de © 2014 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
G E N E T I K U N D E R N Ä H R U N G | I M FO KU S
© 2014 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.biuz.de 1/2014 (44) | Biol. Unserer Zeit | 57
Glucose
Blutgefäß
enteroendokrineZelle
2) Aktivierung der Glucose-Rezeptoren
Glucose
Rezeptor
aktiviertes Signalprotein(Gustducin)
absorptiveEnterocyte
EpithelzellendesDünndarms
3) Sekretion eines Hormon-Cocktails
4) Verstärkung der SGLT1-Transkription
6) Aufnahme von Glucose-Molekülen
5) Einbau der Glucose-Transporter in dieMembran der Mikrovilli
1) Glucose-Moleküle erreichen den Dünndarm
SGLT1
Aus Gründen der Übersichtlichkeit sinddie enterischen Neurone, die das Hormon-Signal von den enteroendokrinen Zellenempfangen (3) und ihrerseits die absorptivenEnterocyten aktivieren (4), nicht eingezeichnet.
ABB. 6 A K T I V I E R U N G D E R S G LT 1 - S Y N T H E S E D U RC H G LU COS E
58 | Biol. Unserer Zeit | 1/2014 (44) www.biuz.de © 2014 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
StRE
DR1
LXRE
PPRE
Gene für Lipid-Transport, Fettsäure-Oxidation, Thermogenese
Gene für Produktionvon Gallensäure
Gene für Produktion von Gallensäure,Lipoproteine, Transferrin und viele andere
Gen-Variante 1c für Fettsäure-, Triglycerid- und Phospholipid-Synthesebzw. Gen-Varianten 1a und 2 für Cholesterin-Synthese
PPAR
langkettige,gesättigte
und ungesättigteFettsäuren
Cholesterinderivate,ungesättigte
Fettsäuren
LXR
gesättigte undungesättigte
Acyl-CoA
HNF4α
gesättigt ungesättigt
langkettige, gesättigte
und ungesättigteFettsäuren
SREBP
gesättigt ungesättigtCH3
CH3
CH3
CH3
HO
CH3
CH3
CH3
CH3
CH3
HO
CH3
CH3
CH3
CH3
CH3
HO
CH3
ABB. 7 F E T T S Ä U R E N A L S G E N - R EG U L ATO R

G E N E T I K U N D E R N Ä H R U N G | I M FO KU S
these der Gallensäuren und solche, die am Glucose-Stoffwechsel beteiligt sind.
SREBP kommt in drei Varianten vor; SREBP-1a und -1c, die Isoformen desselben Gens sind, und SREBP-2.SREBP-1a und -2 aktivieren Gene, die an der Choleste-rin-Synthese beteiligt sind, während SREBP-1c bevor-zugt Gene aktiviert, die an der Fettsäure-, Triglycerid-und Phospholipid-Synthese beteiligt sind. Fettsäurenund Cholesterine beeinflussen diesen Transkriptions-faktor indirekt durch die Blockierung eines Reifungs-schrittes. SREBP wird als inaktive Vorstufe gebildet, vonder erst ein kleiner Abschnitt abgetrennt werden muss,damit das Protein voll funktionsfähig ist. Und Ungesät-tigte Fettsäuren können eben diesen Reifungsschrittund somit die Synthese von Fettsäuren und Cholesterinverhindern.
Züchtungen: Der Einfluss des Menschen auf seine Nahrung
Keine Art nimmt auf die Genome seiner Nahrungsorga-nismen einen derart massiven Einfluss wie Homo sa-piens. Mit Beginn der Neolithischen Revolution vor ca. 10.000 Jahren begannen die Menschen an verschie-denen Orten der Welt, Pflanzen und Tiere von ihren Lebensräumen zu separieren und ihre Vermehrung zukontrollieren. Erwünschte Eigenschaften von Pflanzenwie Größe, Protein-, Zucker- und Stärkegehalt wurdenpositiv selektiert, unerwünschte Eigenschaften wie Bitterstoffe, Säure- und Fettgehalt wurden negativ se-lektiert. Heute müssen die Organismen nicht nur er-nährungsphysiologische Ansprüche erfüllen, sondernimmer häufiger auch den Erfordernissen der verarbei-tenden Industrien genügen.
Beispiel 1: KartoffelnIn den Unterscheidungskriterien der weltweit unge-fähr 5000 Sorten spiegeln sich die verschiedenen Züch-tungsziele wider: Die Form ist für eine industrielle Wei-terverarbeitung entscheidend (lange Formen fürPommes frites und für Chips), eine möglichst einheit-lich gelbe Fleischfarbe wird grundsätzlich von allen be-vorzugt, die Kocheigenschaften sind für die Erzeugungder Endprodukte wichtig (festkochende Sorten alsSpeisekartoffeln, mehligkochende Sorten für Fertig-Rei-bekuchen und Püreeflocken) und ein möglichst hoherGehalt der Stärkeform Amylopektin ist für die Papier-und Klebstoffindustrie entscheidend.Allgemeine Züch-tungsziele sind eine möglichst hohe Lagerfestigkeitund ein möglichst weites Spektrum an Krankheitsresis-tenzen, sowie eine große Schalenfestigkeit, um Verlustedurch Verletzungen während der maschinellen Erntemöglichst gering zu halten. 2011 wurde eine umfang-reiche Analyse veröffentlicht, bei der nicht nur erstmalsdas Genom einer Kartoffelpflanze publiziert, sondernauch das Genom einer ursprünglichen Sorte mit demeiner modernen Sorte verglichen wurde [14]. Gene, die
an der Stärkebildung beteiligt sind, werden bei der mo-dernen Sorte drei- bis achtmal stärker abgelesen als beider Natursorte. Zusätzlich wurde der Transport vonKohlenstoffverbindungen aus den Blättern in die stär-kespeichernden Organellen der Stolone verstärkt. Au-ßerdem werden bei der Kultursorte die Gene für stär-keabbauende Enzyme durchschnittlich 15mal wenigerabgelesen als bei der ursprünglichen Form. Und einkomplexes Regulationssystem, das bei der Wildformden Blütenzeitpunkt an die Tageslänge koppelt, ver-knüpft bei der Kulturform die Tageslänge an die Um-formung der Stolonspitzen zu Knollen, also an die Aus-formung der Kartoffel. Diese und weitere Daten sollenZüchtern nun helfen, schneller und effektiver neue Sor-ten mit definierten Eigenschaften züchten zu können.
Beispiel 2: ApfelsinenDie Kultivierung von Zitrusfrüchten begann vermut-lich vor über 4000 Jahren im südöstlichen Asien und esist heute nicht leicht herauszufinden, welche der vielenArten ursprünglich und welche das Resultat von Züch-tungen und Kreuzungen sind. Von einer vollständigenAufklärung der Verwandtschaftsbeziehungen unter denzahlreichen Zitrusfrüchten ist die Wissenschaft nochweit entfernt, aber ein erster wichtiger Grundstein istnun gelegt. Im Jahr 2013 wurde eine aufwändige Arbeitveröffentlicht, in der der Ursprung der wirtschaftlichwichtigsten Frucht, der Süßorange, beschrieben wird(Abbildung 8) [15]. An welche Stelle des Citrus-Stamm-baums diese einzuordnen ist, war bis dahin völlig un-klar. Sicher war lediglich, dass diese Frucht in Asienschon recht lange kultiviert wird, denn den bislang äl-testen Hinweis auf die Existenz der Süßorange gibt einchinesisches Schriftstück aus dem Jahr 314 v. Chr. Die
© 2014 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim www.biuz.de 1/2014 (44) | Biol. Unserer Zeit | 59
Pampelmuse Mandarine
Süßorange
Mandarine
ABB. 8 Z Ü C H T U N G S W EG D E R S Ü S S O R A N G E

Forschergruppe nahm an, dass die Apfelsine mit denbeiden deutlich älteren Arten Mandarine und Pampel-muse verwandt ist. Das aus den Genomanalysen und Se-quenzvergleichen errechnete Verwandtschaftsverhält-nis der Apfelsine zur Pampelmuse und zur Mandarineergab das irritierende Verhältnis von 1:3, was bedeutet,dass die Apfelsine um den Faktor 3 mehr mit der Man-darine als mit der Pampelmuse verwandt ist. Wäre dieApfelsine eine normale Kreuzung aus Pampelmuse undMandarine, dann hätte das Verwandtschaftsverhältnis1:1 betragen müssen. Untersuchungen und Vergleicheder Chloroplasten der drei Pflanzen brachten die Auf-klärung. Als die Organellen der Apfelsine, der Manda-rine und der Pampelmuse miteinander verglichen wur-den, stellte sich heraus, dass sich die Chloroplasten derSüßorange und der Pampelmuse sehr stark ähnelten.Da Chloroplasten ausschließlich über die Eizellen ver-erbt werden, konnte rekonstruiert werden, dass die „Erfinder“ der Apfelsine die Pampelmuse als Mutter-pflanze und die Mandarine als Vaterpflanze ausgewählthaben mussten. Das ungewöhnliche Verwandtschafts-verhältnis von 1:3 erklärt sich, wenn die weiblichenPflanzen der ersten Hybridgeneration erneut mit derMandarine gekreuzt werden: Süßorange = (Pampel-muse x Mandarine) x Mandarine.
ZusammenfassungZwischen Menschen und ihrer Nahrung bestehen zahlreicheWechselwirkungen und Abhängigkeiten, die sich besondersin Veränderungen der genetischen Ausstattungen und inModifikationen der Genregulationen widerspiegeln. Vier In-teraktionswege konnten identifiziert werden: Das individu-elle Genrepertoire des Menschen beeinflusst die Reaktionenseines Körpers auf die zugeführte Nahrung (Nutrigenetik);Nahrungsbestandteile beeinflussen die Regulation einerVielzahl der Gene (Nutrigenomik); traditionelle Ernährungs-gewohnheiten können zu wechselseitigen genetischen Ver-änderungen der Menschen und seiner Nahrungsorganismenführen (Coevolution); auf Grund der positiven Selektion er-wünschter und der negativen Selektion unerwünschter Ei-genschaften verändert der Mensch das Genrepertoire na-hezu aller seiner Nahrungsorganismen (Züchtung und Kul-tivierung).
SummaryHow humans and their food affect each otherThere are numerous interactions and dependencies be-tween humans and their food. They are reflected especiallyin changes in the genetic compositions and in modificationsof gene regulations. Four pathways of interaction have beenidentified so far: The individual gene repertoire of man de-termines the reactions of his body to the consumed food(nutrigenetics); nutrients are influencing the regulation of avariety of our genes (nutrigenomics); traditional dietaryhabits can lead to reciprocal genetic changes in humansand their food organisms (coevolutional processes); hu-
mans are changing the gene repertorie of almost all of theirfood organisms by positive selection of desired propertiesand by negative selections of unwanted properties (breed-ing and cultivation).
SchlagworteNutrigenetik, Nutrigenomik, Coevolution, Züchtung
Literatur[1] D. Corella et al., Association between the APOA2 promoter poly-
morphism and body-weight in mediterranean and asian populati-ons: replication of a gene-saturated fat interaction, InternationalJournal of Obesity 2011, 35, 666–675.
[2] S.-J. Yin et al., Pharmacogenetic determinants of alcohol metabo-lism and alcoholism in the human alcohol dehydrogenase family,in: Enzymology and Molecular Biology of Carbonyl Metabolism 12(Hrsg.: H. Weiner et al.), Purdue University Press 2006, 161–171.
[3] N. C. E. Duranceaux et al., Ethnic differences in level of response toalcohol between cheinese americans and korean americans, J StudAlcohol Drugs 2008, 69(2), 227–234.
[4] V. Douard, R. P. Ferraris, Regulation of fructose transporter GLUT5in health and disease, Am J Physiol Endocrinol Metab 2008, 295,E227–E237.
[5] G. H. Perry et al., Diet and the evolution of human amylase genecopy number variation, Nature Genetics 2007, 39(10), 1256–1260.
[6] A. Revedin et al., Thirty thousand-year-old evidence of plant foodprocessing, PNAS 2010, 107(44), 18815–19.
[7] F. Höffeler, Geschichte und Evolution der Lactose(in)toleranz, Bio-logie in unserer Zeit 2009, 39(6), 378–387.
[8] R. H. Lewinsky et al., T-13910 DNA variant associated with lactasepersistence interacts with Oct-1 and stimulates lactase promoteractivity in vitro, Hauman Molecular Genetics, 2005, 14(24),3945–3953.
[9] R. F. Margolskee et al., T1R3 and gustducin in gut sense sugars toregulate expression of Na+-glucose cotransporter 1, PNAS 2007,104(38), 15075–80.
[10] V. Gorboulev et al., Na+-D-Glucose cotransporter SGLT1 is pivotalfor intestinal glucose absorption and glucose-dependent incretinsecretion, Diabetes 2012, 61 (1), 187–196.
[11] L. Ma et al., ChREBP*Mlx is the principal mediator of glucose-indu-ced gene expression in the liver, J Biol Chem 2006, 281(39),28721–30.
[12] St. D. Clarke, Polyunsaturated fatty acid regulation of gene tran-scription: a molecular mechanism to improve the metabolic syn-drome, Journal of Nutrition 2001, 131(4), 1129–1132.
[13] E. Duplus et al., Fatty acid regulation of gene transcription, Journalof Biological Chemistry, 2000, 275(40), 30749–52.
[14] Potato Genome Sequencing Consortium, Genome sequence andanalysis of the tuber crop potato, Nature 2011, 475, 189–197.
[15] Q. Xu et al., The draft genome of sweet orange (Citrus sinensis), Nature Genetics 2013, 45(1), 59-68.
Der AutorFritz Höffeler ist Diplombiologe und studierte zu-sätzlich Erziehungswissenschaft mit dem Schwer-punkt Didaktik. Nach einigen Jahren der mikrobio-logischen Forschung arbeitet er als Wissenschafts-Autor, -Illustrator und -Berater. 1999 gründete erArt For Science, ein Unternehmen, das Wissen-schaft durch Wort, Schrift und Bild vermittelt. ZurGenetik der Ernährung hat er unlängst ein Buchveröffentlicht, das im Hirzel Verlag erschienen ist.
60 | Biol. Unserer Zeit | 1/2014 (44) www.biuz.de © 2014 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim
Korrespondenz: Fritz HöffelerArt For ScienceStellinger Steindamm 4822527 HamburgE-Mail: [email protected]:www.Art4Science.de





























